Отбор лучших (плюсовых) деревьев
МЕТОДЫ ЛЕСНОЙ СЕЛЕКЦИИ. ОТБОР
Результативность селекции в первую очередь зависит от применяемых методов. К настоящему времени наиболее распространенными являются методы отбора и гибридизации. Кроме того, в последние годы активные работы ведутся по разработке методов полиплоидии, мутагенеза, культуры изолированных клеток и тканей, клеточной и генной инженерии и др.
Виды отбора
При всех селекционных работах отбор является наиболее важным и решающим процессом. В его учение значительный вклад внесли Ч. Дарвин, С. Райт и И.И. Шмальгаузен. Различают естественный и искусственный отбор.
Естественный отбор. Этот вид отбора происходит под действием климата (тепло, влага, свет, ветер), почвенных условий (мехсостав, физические, химические свойства, трофность и др.), влияния рельефа и живых организмов (паразитизм, конкуренция и др.). Естественный отбор искореняет все неприспособленные к данным условиям произрастания биотипы, оставляя только адаптировавшиеся расы или популяции, которые вовлекаются в селекционный процесс, т.е. естественный отбор — это выживание более приспособленных организмов в борьбе за жизнь. В результате естественного отбора сохраняются любые жизненно важные признаки, действующие на пользу организма и вида в целом, и образуются новые формы и виды. Естественный отбор встречается в форме центростремительного (стабилизирующего), центробежного (дизруптивного) и линейного (направленного) отборов, а также в форме комбинаций этих видов отбора (И.И. Шмальгаузен, 1946; Н.П.Дубинин, 1986).
Центростремительный (стабилизирующий) отбор — это отбор, при котором в случае сохранения среднего уровня условий среды обитания при репродукции сохраняются особи, выражение комплекса признаков у которых приближается к среднему для всей популяции. Отклоняющиеся от модального типа особи элиминируются. Этот отбор сохраняет в популяции определенную однородность особей, то есть стабилизирует популяцию.
Центробежный (дизруптивный, разрывающий) отбор — это естественный отбор, который осуществляется в том случае, когда исходная популяция является настолько неприспособленной к имеющимся условиям внешней среды, что практически любое отклонение особи от модального для популяции типа приобретает селекционные преимущества и поддерживается отбором в процессе репродукции. При дизруптивном отборе исходная популяция расчленяется на ряд менее объемных, но более узколокально приспособленных популяций.
Линейный (направленный, движущий) отбор — это естественный отбор, когда при размножении преимущество получают формы с уклонением признаков в определенном направлении от среднего для популяции типа. В этом же направлении в процессе непрерывного линейного отбора сдвигается и модальный тип популяции.
Искусственный отбор. Этот вид отбора проводится человеком. В результате этого отбора на основе наследственности и изменчивости создаются новые хозяйственно ценные формы и сорта, а также поддерживаются (сохраняются) признаки сорта в процессе его семеноводства. Различают бессознательный и сознательный (методический) искусственный отбор.
Бессознательный отбор — это наиболее ранняя форма селекции, при которой человек сохранял лучшие и уничтожал худшие формы. При этом цель создания новых форм и сортов им не ставилась, и результаты отбора были непредвидимыми.
Методический отбор — это сознательно применяемый человеком метод селекции, при котором заранее ставится цель создания сортов растений с нужными признаками и свойствами. В основном методический отбор проводится по комплексу признаков, формирующих нужную для селекционера форму, и лишь в специальных случаях — по одному какому-нибудь признаку (односторонний отбор), но это, как правило, приводит к снижению общей жизнеспособности организма.
Выделяют прямой и непрямой (косвенный отбор). Прямой отбор ведут по селективному признаку. Непрямой — по косвенному, который связан каким-либо образом с прямым. В пределах онтогенеза проводят так называемую раннюю диагностику развития признака, то есть пытаются оценить свойства селектируемых образцов на ранних этапах онтогенеза.
Методический отбор делится на массовый и индивидуальный.
Массовый отбор
2.2.1. Теория массового отбора и возможность его использования в лесном хозяйстве
При массовом отборе выделяют множество индивидов, наиболее отвечающих задачам селекции, и размножают их совместно. В основе массового отбора лежит оценка фенотипа. Стихийно он применялся человеком с незапамятных времен, затем сознательно использовался при выведении многих высокопродуктивных сортов культурных растений и пород домашних животных. Он не утратил своего значения и в настоящее время, так как он наиболее прост и доступен, а также может нередко давать хорошие результаты.
Однако этот тип отбора имеет и существенные недостатки, обусловленные тем, что по фенотипу трудно однозначно судить о генотипе отбираемых особей, от которого зависит эффективность отбора. Фенотип представляет собой результат взаимодействия генотипа и среды, а на большинство количественных признаков, интересующих селекционера, влияние последней весьма значительно. Кроме того, в фенотипе не проявляются рецессивные гены, находящиеся в гетерозиготном состоянии, но способные проявиться в потомстве. Ввиду этого массовый отбор, как правило, действует очень медленно, а иногда оказывается и вовсе безрезультатным. Последнее характерно главным образом для тех случаев, когда популяция, в которой ведется отбор, гомозиготная или почти гомозиготная по генам, определяющим отбираемый признак, то есть коэффициент его наследуемости близок к нулю.
Хорошей иллюстрацией безуспешности отбора в гомозиготных популяциях, несмотря на наблюдаемую в них фенотипическую изменчивость, могут служить опыты Иоганнсена, проведенные еще в началеXX века. Он изучал изменчивость веса семян фасоли и действие отбора на этот признак. Семена в смеси, собранной с разных растений изучаемого им сорта, характеризовались значительной изменчивостью массы — приблизительно от 0,25 до 0,75 г (средняя масса около 0,5 г). Поскольку нормально фасоль размножается самоопылением, то все семена в этой смеси должны были быть гомозиготными. Однако генотипически они могли быть различными, поскольку происходили от разных родителей. То и другое подтвердилось результатами последующей работы.
Из исходной смеси были отобраны и высажены семена разной массы. Растения, выращенные из наиболее тяжелых семян, дали тоже тяжелые семена (средняя масса около 0,66 г), а растения, выращенные из легких семян, произвели легкие семена (средняя масса около 0,37 г). Т.е. отбор здесь был эффективным. Однако дальнейший отбор самых тяжелых и самых легких семян на протяжении шести поколений в пределах отдельных выделенных таким образом чистых линий оказался совершенно безрезультатным, хотя в каждой линии наблюдалась некоторая изменчивость массы семян. Следовательно, в гомозиготном материале отбор бессилен, так как там изменчивость зависит не от генетических, а от паратипических факторов. Чистые линии остаются константными до тех пор, пока их гомозиготность не будет нарушена возникшей мутацией. При проведении массового отбора по фенотипу в гетерозиготной популяции перекрестников изменение признака в желаемую сторону обычно идет относительно быстро в первых поколениях, а затем замедляется по мере того как отбираемые особи становятся все более гомозиготными по желательным генам. Однако иногда отбор по фенотипу остается эффективным в течение десятков поколений, как это бывает при высокой генетической гетерогенности популяций и большом числе отбираемых в каждом поколении особей, что препятствует наступлению гомозиготности (С.М. Гершензон, 1979).
Учитывая значительную гетерогенность лесных древесных пород и трудность применения к ним других видов отбора, массовый отбор широко используется в их селекции. Выбор желательных, наиболее пригодных для селекции типов (как правило, небольшой процент), называют позитивным массовым отбором, а искоренение нежелательных форм — негативным. Последний, не следует смешивать с отрицательно действующим отбором, когда из популяции удаляются наследственно ценные компоненты насаждения и сохраняются менее ценные (например, в результате так называемых выборочных или приисковых рубок).
К негативному массовому отбору в лесных популяциях относятся: рубки ухода, когда в первую очередь удаляют наименее ценные, больные и худшие растения; сплошная рубка насаждений, не соответствующих местообитанию для предотвращения опасности распространения непригодных наследственных свойств через пыльцу; отбор в питомнике, когда удаляют нежелательные типы по ветвлению и устойчивости среди сеянцев и растений.
При позитивном массовом отборе в лесном хозяйстве применяют разные методы:
• отбор лучших географических происхождений, или климатипов;
• отбор лучших эдафотипов в пределах какого-либо географического района;
• отбор лучших (плюсовых) насаждений;
• отбор лучших (плюсовых) деревьев;
• отбор лучших сеянцев в питомниках и семян по крупности и качеству.
Ниже дана краткая характеристика этих видов массового отбора.
2.2.2. Отбор географических происхождений, или климатипов
Изучение насаждений из привозных семян еще в XIX веке показало, что лучшим ростом и качеством отличаются насаждения, созданные из местных семян. Это было замечено при выращивании ели на Тимирязевской даче из семян, привезенных из Германии (Н.С. Нестеров, 1935, цит. по А.В. Альбенскому, 1959). В Воронежской области в культурах сосны, созданных из семян, импортированных из Германии (г. Дармштадт), вырастали большей частью так называемые “пьяные” деревья (М.М. Вересин, 1981). То же явление было отмечено и у других пород, например, при выращивании ольхи черной в Германии из семян, привезенных из Бельгии, когда первоначально быстрый рост сменялся его притуплением и заболеванием деревьев в целом.
Исключением из этого правила является южная Швеция, где насаждения ели из семян, привезенных из Средней Европы, растут лучше местных. Это объясняется тем, что после ледникового периода ель не могла проникнуть в южную Швецию с юга через Балтийское море, она постепенно продвигалась туда из северной Швеции через Финляндию, подвергаясь отбору суровым климатом. Наиболее быстрорастущие, но чувствительные к холоду биотипы, вероятно, выпали. В результате этого ели из южной Швеции растут медленнее привозных (Э. Ромедер, Г. Шенбах, 1962). Имеются и другие исключения, когда интродуцированные географические экотипы растут лучше местных (М. Giertych, 1991).
Для изучения влияния географического происхождения или “климатических экотипов” на рост, развитие, устойчивость и качество древостоев уже в начале XIX века были заложены первые опыты Луи-де-Вильмореном во Франции (1823-1832 гг.), М. К. Турским в России (1877-1891 гг.), Кинитцем в Германии (1877-1878 гг.), Цизляром в Австрии (1893-1896 гг.), Энглером в Швейцарии (1890), Шотте в Швеции (1903-1904 гг.), В.Д. Огиевскимв Европейской части России (1910-1916 гг.) и др. Проанализировав результаты этих и других опытов, О.Г. Каппер (1946) впервые у нас в стране дал полную картину характера поведения климатических экотипов. Учитывая значение географического происхождения семян для лесного хозяйства, приведем хотя бы в сокращенном виде эти выводы, поскольку на них базировались все последующие исследования географических экотипов, по крайней мере, в нашей стране.
1. Общие выводы:
• Главные лесные породы различного географического происхождения имеют ясно выраженные наследственные свойства по габитусу крон, ветвей и других морфолого-физиологических признаков. Это показали еще опыты Цизляра с силезской и тирольской лиственницами, Энглера и Мюнхас елью и др.
• Переброска семян из одного климатического района в другой должна осуществляться на основании опытов по изучению климатических экотипов.
• Лучший производственный эффект, наибольшую успешность и благонадежность имеют, как правило, посевы в посадках из местных семян.
2. Биологические особенности климатипов:
• Фенология отдельных климатипов имеет свои особенности. Энглерв Швейцарии отмечал, что в 4-5-летнем возрасте разница во времени окончания роста у скандинавских и русских сосен, по сравнению со швейцарскими и южно-германскими, равна двум неделям, а в 6-7-летнем возрасте эта разница доходит до трех недель. В общем, начало вегетации разных климатипов зависит от хода весны. При наступлении сразу высоких температур вегетация начинается почти у всех климатипов (одновременно), тогда как медленное нарастание высоких температур весной вызывает развитие северных и южных форм в разное время. Окончание вегетации происходит у разных климатипов не одновременно. Более северные климатипы заканчивают вегетацию раньше, а южные — позже.
• Отношение к крайностям температуры у отдельных климатипов различно. Шотте в Швеции отмечал большую повреждаемость морозом, а также большую искривленность стволов от навала снега у южных форм сосен по отношению к местным соснам. В.Д. Огиевский наблюдал большую устойчивость северных сосен в условиях Ленинградской области по сравнению с южными формами. Цизляр пришел к заключению, что перенос черешчатого дуба из умеренного климата в более теплый создает морозостойкие культуры. В то же время перенос дубов из теплой области в более умеренный климат усиливает опасность повреждения морозом дубовых культур. В большей части России однолетние сеянцы сосны с наступлением морозов принимают темно-фиолетовый оттенок хвои, что делает их более устойчивыми к морозам, тогда как западноевропейские формы сохраняют ярко-зеленую окраску и нередко побиваются морозами.
• Репродуктивная способность климатипов тоже различна. У северных форм при произрастании их в более южных условиях возраст возмужалости наступает раньше, причем количество шишек у северных форм меньше, чем у местных.
• Северные формы сосны обыкновенной с более выраженным эпидермисом на хвое меньше повреждаются шютте, чем южные формы.
• Семена северной сосны сохраняют свою всхожесть более продолжительное время, чем семена южной.
• Требовательность к свету у климатипов различна. Для отдельных климатипов существует свой оптимум освещения. Так, в условиях Московской области, по Н.С. Нестерову, вологодская сосна менее требовательна к свету, чем владимирская.
• Северные климатипы более устойчивы к навалу снега из-за наличия у них более узких крон, что связано еще и с условиями освещения на севере.
• Дифференциация стволов с возрастом наблюдается в связи с разным отношением к свету, к заморозкам, а также с разной формой кроны и другими свойствами.
3. Изменчивость морфологических признаков у климатипов:
• Форма кроны у деревьев северных климатипов узкая,частопирамидальная, тогда как у южных форм она более широкая и раскидистая.
• Длина хвои и листьев у северных форм более короткая, чем у южных форм.
• Размеры шишек и длина семян у северных климатипов меньше, чем у южных.
• Цвет семян северных климатипов более светлый, чем у южных форм.
• Более быстрый рост соответствует большей величине листьев.
4. Анатомическая изменчивость климатипов:
• Мощность покровных тканей — эпидермиса и гиподермы — в хвое сосны к северу увеличивается. По другим анатомическим признакам определенные тенденции О.Г. Каппер не выявил.
5. Биохимические исследования климатипов:
• Содержание смол увеличивается от северныхк южным климатипам.
• Содержание жиров уменьшается с севера на юг.
6. Анализ роста климатипов:
• Местные растения (в среднем) имеют наибольшие показатели прироста по высоте.
• Дубы, полученные от быстрорастущих материнских деревьев, обнаруживают быстрый рост в высоту, если климат родины и климат места разведения не сильно отличаются один от другого.
• Рост культур (в общем) тем более медленный, чем географически дальше на север, восток или юго-восток отстоят районы заготовок семян от района культур.
• Перенос южных сосен в северные условия, в связи с длительной их вегетацией, приводит к уменьшению процента поздней древесины, а следовательно, и к большей рыхлости и ломкости ветвей и деревьев.
• Коэффициенты формы сосны (q2) увеличиваются от центральных районов на север и северо-восток и уменьшаются на запад, юго-запад и юго-восток.
• Кривизна стволов сосны является признаком наследственного порядка и в то же время может иметь место при нарушении нормальных жизненных условий в результате переноса сосны из одних климатических условий в другие.
Однако О.Г. Каппер не ограничился этими выводами. Он проанализировал методы и условия закладки этих опытов и нашел целый ряд недочетов, которые предложил устранить в последующих работах. В частности, он обращал внимание на необходимость учета не только географических координат, но и экологических условий произрастания насаждений, из которых собираются семена, а также фиксации фенологических форм и даже отдельных деревьев. Он был категорически против составления опытных образцов путем перемешивания семян из разных популяций. Многие из его рекомендаций были учтены при закладке географических культур более позднего времени (М.М. Вересин, 1963; Р.И. Дерюжкин, 1969; А.И. Ирошников, 1977; A.M. Шутяев, 1998 и др.). Накопленный опыт позволил определить оптимальные расстояния по переброске семян тех или иных лесных древесных пород. Усредненные данные, полученные в этих работах, колебались, например, для сосны, по М.М. Вересину (1963), от 300 до 600 км при переброске семян с севера на юг. При переброске их в обратном направлении расстояние уменьшается вдвое. При перемещении семян с востока на запад эти расстояния составляют 500-850 км, в обратном направлении тоже в два раза меньше. В горных районах допускается перемещение семян сверху вниз до 400 м; снизу вверх расстояние уменьшается в два раза.
Более детально опыт по перемещению семян был обобщен в специальном издании “Лесосеменное районирование основных лесообразующих пород в СССР” (1982), на котором следует остановиться более подробно. В основу его, по информации разработчиков, положены следующие данные: результаты исследований внутривидовой изменчивости древесных пород в географических культурах и природных популяциях; материалы по изучению истории расселения видов в межледниковые периоды, численности популяции в разных частях ареала; наличие современных изоляционных барьеров; результаты исследования ареалов других видов растений-эдификаторов; особенности плодоношения в многолетнем цикле и качество семян; интегральная оценка природно-климатических факторов; физико-географическое, агроклиматическое, почвенное, геоботаническое, геоморфологическое, лесорастительное и лесохозяйственное районирование.
Кроме того, при его разработке использованы результаты исследований географических культур (семенных потомств географически отдаленных популяций) и рекомендации по переброске семян и лесосеменному районированию М.К. Турского, А.Н. Соболева, Н.С. Нестерова, В.Д. Огиевского, В.Н. Сукачева, С.А. Самофала, А.П. Тольского, А.И. Колесникова, В.В. Гурского, А.Б. Жукова, П.К. Фальков-ского, Ф.И. Фомина, В.М. Ровского, В.М. Обновленского, О.Г. Каппера, М.М. Вересина, А.С. Яблокова, Л.Ф. Правдина, В.П. Тимофеева, С.А. Ростовцева, Е.И. Еньковой и др. Районирование описано раздельно по породам, что связано с различиями в географическом распространении, экологическими особенностями и характером внутривидовой изменчивости главных лесообразующих пород.
За основную единицу лесосеменного районирования принят лесосеменной район, т.е. некоторая территория (в пределах ареала вида) со сравнительно однородными природными условиями и генотипическим составом популяций, с ясно выраженными природными и лесохозяйственными особенностями, которые обусловливают общий характер основных мероприятий по организации лесного семеноводства и использованию однородного по наследственным свойствам семенного материала. Лесосеменной район в ряде случаев разделяется на несколько подрайонов, т.е. на территории, характеризующиеся большей однородностью лесорастительных условий и генотипического состава популяций. Лесосеменные районы расположены на территории одного или нескольких административных подразделений (республика, край, область) или их части.
В пределах ареала вида лесосеменные районы не равноценны между собой по площади, представительству отдельных лесных формаций, перспективам развития семенной базы, лесокультурному фонду данной породы и т.п. Схемы лесосеменного районирования некоторых основных лесообразующих пород нашей страны, взятые из этого документа, приведены на рисунках 2. — 2. .
Для каждого лесосеменного района (подрайона) предусмотрено использование семян из популяций определенного эколого-геогра-фического происхождения. Предпочтение отдается семенам местных и смежных с ними популяций, наиболее адаптированных к природным условиям района. Под местными понимаются семена, собранные непосредственно в пределах лесосеменного района. Семена, заготовленные в других лесосеменных районах, называются инорайонными.
В равнинных областях, отличающихся постепенным изменением биологических признаков деревьев, допускается переброска семян по территории всего лесосеменного района, а также между контактирующими районами и подрайонами. В горных районах, характеризующихся резким изменением условий местопроизрастания в зависимости от высоты местности, перемещение семян рекомендуется лишь в пределах 200-400 м по вертикали от места их заготовки.
В каждом лесосеменном районе (подрайоне) семена предложено собирать раздельно по хозяйственным группам типов леса. В ценных массивах в семенные годы должен обеспечиваться полный сбор семян для последующего использования их в годы неурожая. Для древесных пород с отчетливо выделяющимися фенологическими формами семена заготавливают и используют раздельно по этим формам.
В целом можно отметить, что в цитируемом документе отражен весь опыт, накопленный к тому времени в стране в данном направлении исследований. Указанное лесосеменное районирование было разработано по заданию Государственного комитета СССР по лесному хозяйству (приказ №29 от 6 февраля 1973) отраслевыми и академическими институтами лесного хозяйства и высшими учебными заведениям. Проект сводного лесосеменного районирования скоординирован и подготовлен: Всесоюзным научно-исследовательским институтом лесоводства и механизации лесного хозяйства (Е.П. Проказиными Б.Н. Куракиным) — по сосне и ели в европейской части СССР; Институтом леса и древесины им. В.Н. Сукачева СО АН СССР (А.И. Ирошниковым) — по сосне и ели в азиатской части СССР, а также по сосне кедровой сибирской, лиственнице и пихте сибирской; Центральным научно-исследовательским институтом лесной генетики и селекции (A.M. Шутяевым) — по дубу;
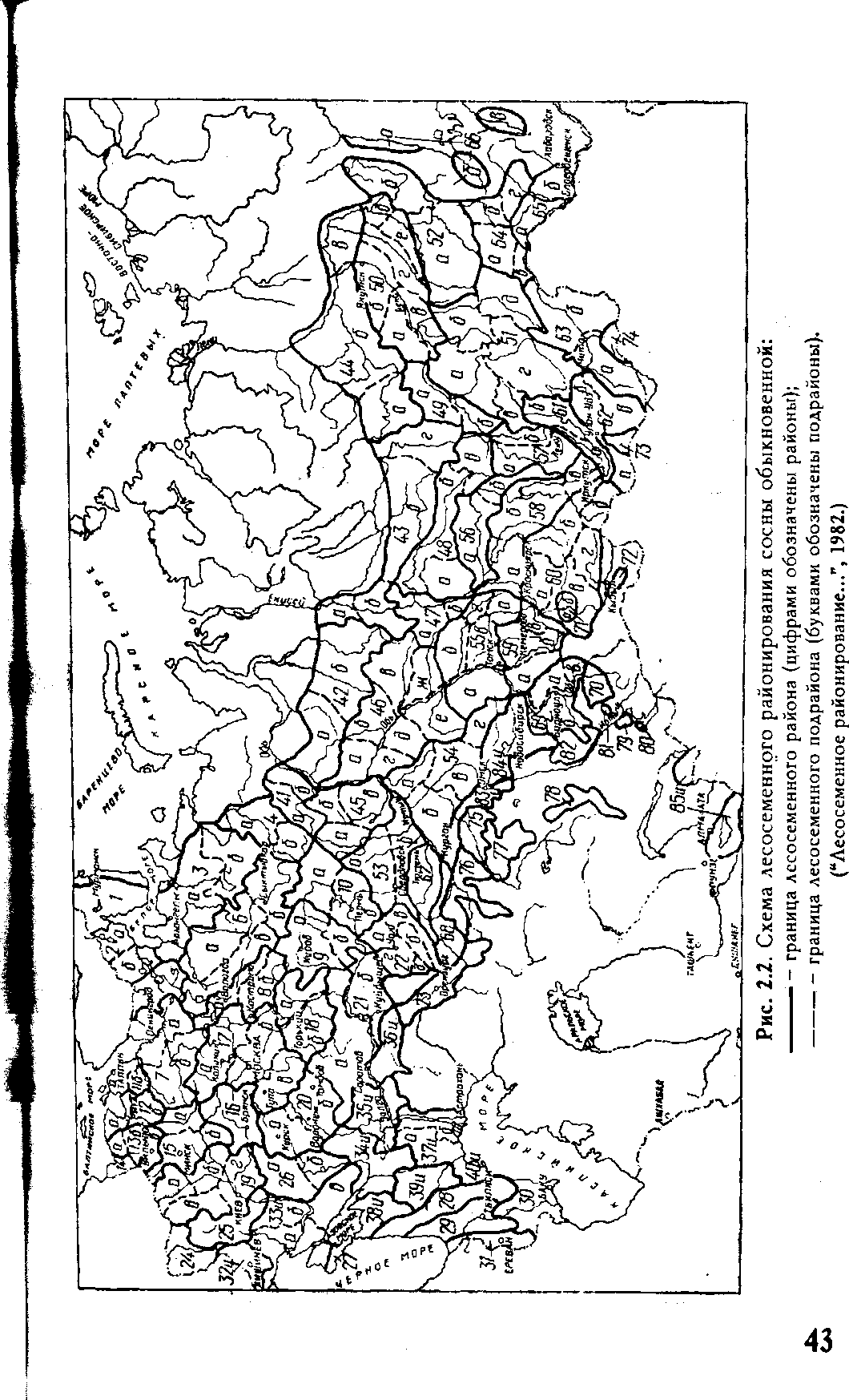
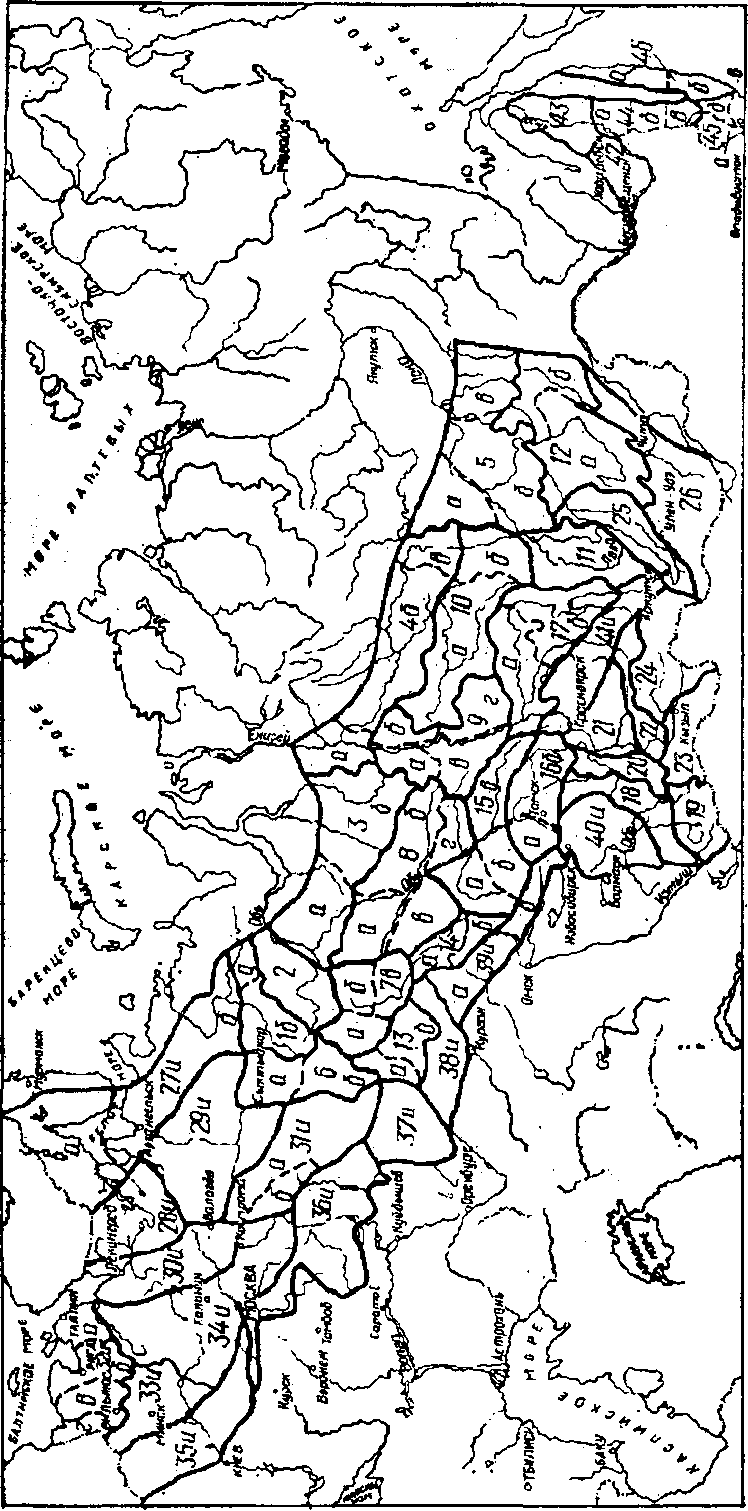
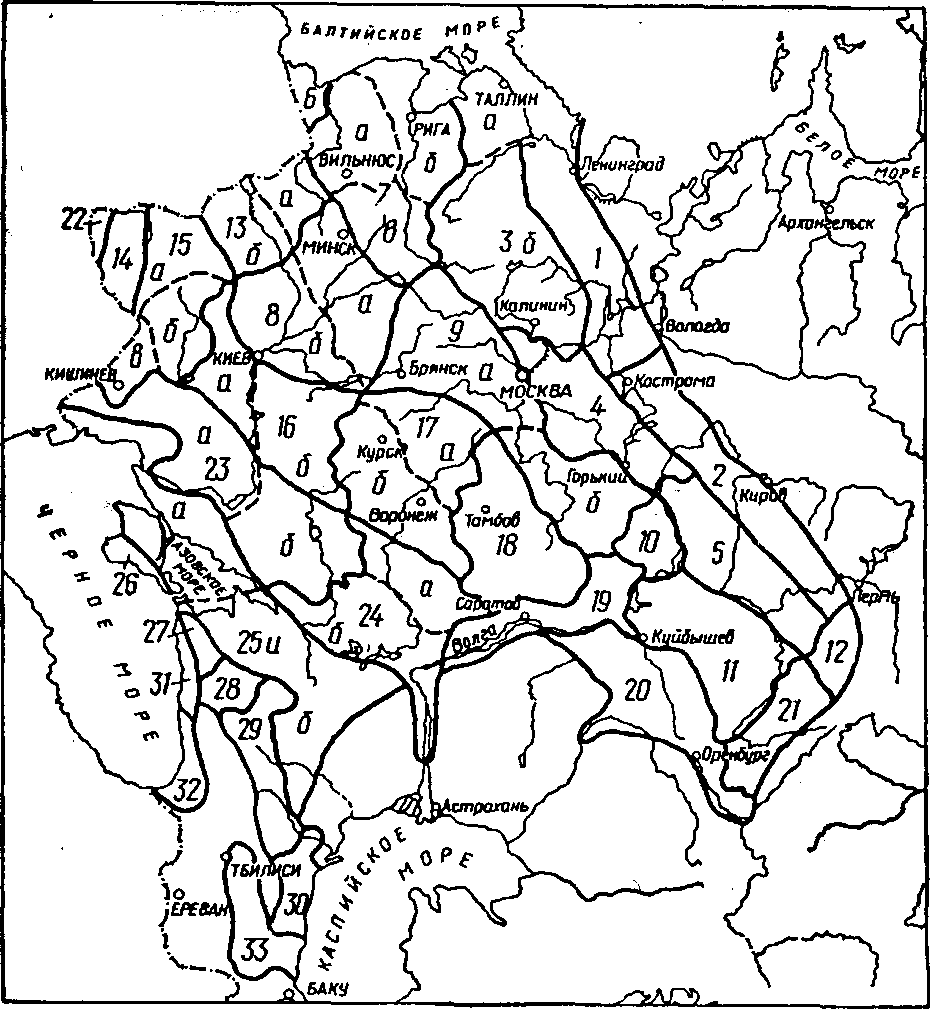
Рис.Схема лесосеменного районирования дуба черешчатого:
—— — граница лесосеменного района (цифрами обозначены районы);
•—— — граница лесосеменного подрайона (буквами обозначены подрайоны). ("Лесосеменное районированию...", 1982.)
Тбилисским институтом леса (К.Д. Нацвлишвили) — по пихте кавказской; Украинским научно-исследовательским институтом лесного хозяйства и агролесомелиорации (П.И. Молотковым) — по пихте европейской и буку; Среднеазиатским научно-исследовательским институтом лесного хозяйства (М.Я. Парфеновым) — по саксаулу.
Перечень исследователей, которые участвовали в разработке этого лесосеменного районирования по отдельным породам, насчитывает десятки фамилий. Их список приведен выше цитируемого сводного документа. Необходимо также отметить активную позицию и существенный вклад в организацию всего этого огромного дела работников тогдашнего Госкомлеса А.И. Новосельцевой и И.Н. Чеботарева.
Учитывать влияние географического происхождения семян на рост насаждений важно не только для автохтонных пород, но в не меньшей степени и для интродуцированных пород. Так, исследование роста различных происхождений сосны скрученной (Pinus contortа) из Канады в Швеции показало, что различные происхождения в разных регионах Швеции показывают разный результат. При этом разница может достигать 60-70 м3га по запасу в 18-летнем возрасте (К. Lindgren, 1993,цит. по Н. Hattemer, F. Bergmann, М. Ziehe, 1993). Следовательно, и при интродукции необходимо прежде всего определять наиболее пригодные части ареала интродуцируемой породы, перемещение репродуктивного материала из которых даст наибольший хозяйственный эффект. Этот аспект также был отражен в отечественном лесосеменном районировании.
В рассмотренном выше сводном документе было подготовлено интродукционное районирование основных лесообразующих пород:
а) в европейской части страны по четырем породам: лиственнице — Р.И. Дерюжкиным; ели — Б.Н. Куракиным, A.M. Шутяевым; сосне сибирской кедровой — М.В. Твеленевым, В. И. Некрасовым; сосне обыкновенной — И.Н. Патлаем; б) в азиатской части по интродукции основных лесообразующих пород А.И. Ирошниковым и В.И. Мосиным и др.
Рассмотренный документ был обязательным для всех государственных предприятий и организаций, осуществляющих ведение лесного хозяйства независимо от их ведомственной принадлежности. Все прежние документы утрачивали силу. Однако с течением времени, и этот документ потребовал уточнений в части как большей детализации географического происхождения, так и более тщательного учета почвенно-экологических условий.
2.2.3. Отбор лучших эдафотипов
Закономерности, отмеченные в поведении климатипов, не всегда проявлялись в результате того, что при сборе семян не учитывались почвенно-экологические условия. На это в свое время обратил внимание О. Г. Каппер (1946), некоторые рекомендации разработал М.М. Вересин (1946), которые вошли и в более поздние справочники и учебники. Основные выводы М.М. Вересина сводились к трем положениям:
1) в целях повышения успешности и продуктивности насаждений сбор и использование семян следует, как правило, производить с учетом типов леса и условий произрастания, группируя их соответствующим образом;
2) в пределах каждой группы сбор семян следует производить в наиболее продуктивных древостоях, на лучших почвах;
3) при нехватке семян в некоторых группах переброску их можно допускать из смежной группы с более продуктивными древостоями и почвами в менее продуктивную в данном экологическом ряду, но не наоборт, а для равноценных по продуктивности групп — с учетом их экологических условий и диапазона приспособляемости получаемого от каждой из них лесокультурного материала.
При этом для дуба в условиях лесостепи он наметил пять групп типов леса и условий произрастания, которые необходимо разграничивать при сборе и использовании желудей.
• Первая группа охватывает нагорные свежие дубравы и переходные от них к влажным и сухим — дубняки снытевый, снытево-осоковый и осоковый (эдатопы D2-3 , D2 ,D2-1).
• Вторая группа — нагорные дубняки на солонцеватых почвах “солонцах” (осолоделые суглинки) (эдатопы D1-0, D0 , С0).
• Третья группа — дубняки по тальвегам и нижним склонам оврагов, на овражном аллювии и делювиальных шлейфах (эдатоп D3).
• Четвертая группа — комплекс пойменных дубрав, эдатопы колеблющегося увлажнения (D2-5,С2-5).
• В пятую группу выделились подборовые левобережные дубняки. Исследование желудей, сеянцев и культур обнаружили наследственную и селекционную неравноценность дубняков указанных групп местопроизрастания. Так, сеянцы из желудей второй группы характеризовались чрезвычайно большим коэффициентом изменчивости по энергии роста, обилием вильчатых и тройчатых растений. Культуры из желудей этой группы в эдатопе D2. показывали худшую приживаемость и в 17-летнем возрасте отставание в росте на 25-30% от культур местного (D2) происхождения.
Сеянцы из желудей третьей группы в вегетационных опытах обнаружили меньшую засухоустойчивость и продуктивность транспи-рации и большую интенсивность последней, чем сеянцы плакорного дуба (первая группа), а наибольшая энергия фотосинтеза наблюдалась у них при более высокой влажности почвы. Крупные желуди дубняков четвертой группы дают крупные сеянцы с малым количеством вильчатых и тройчатых побегов. Изменчивость по энергии роста у них невелика. По данным вегетационных опытов и культур дубки пойменного происхождения значительно менее засухоустойчивы и менее продуктивны в плакорных условиях, чем местные.
Наиболее четко прослеживается своеобразие для подборовых дубняков. Неблагоприятные для дуба почвенные условия и длительное произрастание его во втором ярусе в сосняках предполагают образование здесь эдафо-фитоценотического “суборевого” экотипа дуба с рядом специфических свойств: повышенной изменчивостью, теневыносливостью, более медленным ростом и наклонностью к образованию ветвистых и корявых стволов. Так, однолетние сеянцы из подборовых желудей (эдатопы С2,B2 А1-В1) показали чрезвычайную изменчивость по энергии роста и обилие вильчатых экземпляров, особенно сеянцы из желудей А1-В1 Последние, несмотря на мелкие размеры желудей, давали в посевах-коллекциях наряду с “карликами” единичные самые крупные по высоте сеянцы и наибольшее количество сеянцев с третьим побегом. Эти черты весьма сближают их с сеянцами “солонцового” дуба. Опыты с использованием смешанных суборевых желудей в культурах в эдатопе D2 к 14 годам обнаруживали чрезвычайную пестроту и в среднем худший рост суборевых дубков, чем подроста местного происхождения.
Необходимо также заметить, что наличие двух больших групп наследственно-экологических форм — пойменной (или болотной) и нагорной (или суходольной), отмеченное для дуба, характерно также для ряда других пород: ясеня (Мюнх, П.С. Погребняк), лиственницы даурской (С.Д. Георгиевский), ивы (S. rosmarinifolia; B.H. Сукачев), сосны обыкновенной и сибирской, осины, ели, пихты и других пород.
При нехватке желудей в пределах отдельных групп можно считать условно допустимой переброску их из группы первой во вторую, третью и четвертую, но не наоборот. В отношении низкопродуктивных дубняков (IV и V бонитеты) второй группы необходимы дальнейшие исследования природы их наследственной изменчивости. Возможно, что в ряде случаев окажется целесообразным высевать вместо местных семян семена от лучших древостоев,как эторекомендовал еще Буркгардт для сосны.
В отношении сбора и использования семян сосны для условий лесостепи и области хвойно-широколиственных лесов (европейская часть России) М.М. Вересиным намечено также пять групп типов леса и местопроизрастания.
• Первая группа — сложные сосняки (субори) — эдатопы B2, B3 С2 С3 и B0, B1, С0, С1,. К последней группе могут быть условно причислены и меловые боры.
• Вторая группа — свежие боры: брусничники, верещатники, кисличники, черничники и географически замещающие последние в южных массивах молиниевые травяные боры (эдатопы А2, А3).
• Третья группа — лишайниковые и лишайниково-травяно-степные боры (эдатопы А0, А1, А1-В1).
• Четвертая группа — боры сфагновые и долгомошники (эдатопы А5, А4).
• Пятая группа — травяно-болотно-пойменные сосняки. Преимущественно субори (эдатопы B4,B5,С4), отчасти боры (травяно-болотные).
Группа свежих боров по наследственным свойствам сосны определенно обособляется от сосняков болотных и суборей, менее существенно от группы сухих боров. При нехватке семян можно допустить переброску их из ближайших по экологическому характеру типов местопроизрастания; из группы первой в группы вторую, третью, пятую; из группы второй (свежие и влажные боры) в группу третью (из свежих боров) и четвертую (из влажных боров-черничников и молиниозников). В отношении сухих и болотных боров (группы третья и четвертая) необходимы более обстоятельные исследования природы наследственной изменчивости сосняков.
М.М. Вересин (1946) отмечал, что это ориентировочные схемы. Однако до настоящего времени вопрос о влиянии экологического происхождения семян на рост и устойчивость и в целом на характер создаваемых из них насаждений изучен недостаточно.
2.2.4. Отбор лучших (плюсовых) насаждений
Отбор насаждений вышесреднего качества — это одно из направлений позитивного отбора. У нас в стране его называют еще “групповым” (А.Я. Любавская, 1982; М.М. Котов, 1997), хотя по принципиальным особенностям он является типично массовым отбором. Производится он следующим образом. В данном географическом или лесосеменном районе выбирают лучшие по продуктивности, форме стволов и другим качествам насаждения и только их используют для заготовки семян и последующего выращивания растений.
Степень генетического улучшения следующей генерации зависит в первую очередь от того, насколько возможно по фенотипу отличить высокоценные наследственные качества от малоценных. Нередко хорошие наследственные качества не проявляются из-за плохих условий местопроизрастания, и наоборот, в хороших условиях более высокую ценность могут показывать фенотипы с относительно худшими наследственными качествами.
Однако есть такие признаки, как, например, слабая устойчивость к снеголому равнинных елей и высокая — горных елей, которые четко проявляются в фенотипе (в районе обильных снегопадов). Сбор семян с неповрежденных снеголомом растений может обеспечить значительное улучшение наследственных свойств будущего поколения. В целом результаты этого фенотипического отбора можно оценить уже в течение нескольких десятилетий.
Кроме отбора лучших насаждений, в лесах может проводиться негативный отбор в виде верхового и отборного прореживания. При этом для успешного роста лучших деревьев удаляют менее продуктивные независимо от качества их наследственных задатков. Так как успех этого метода также зависит от возможности распознавания генотипов вышесреднего качества по фенотипу, то больших результатов от этого метода ожидать не приходится. Хотя мероприятия по приданию лесу нужного внешнего вида и повышению его полезных качеств, способствуют некоторому улучшению состава наследственных задатков, накапливающихся в течение многих поколений (Э. Ромедер, Г. Шенбах, 1962).
Отбор лучших (плюсовых) деревьев
Выбирают наиболее соответствующие задачам селекции старые деревья, которые также называют отборными, материнскими, плюсовыми или элитными. Из черенков этих деревьев без предварительной оценки их наследственных свойств создают прививочные плантации (так называемые прививочные семенные плантации первого поколения или порядка). Ожидается некоторое повышение продуктивности насаждений, созданных из семян, полученных на этих плантациях. Однако эти ожидания особенно при селекции на продуктивность по массе не всегда оправдываются. При селекции на другие признаки (форму ствола и кроны, устойчивость и др.) массовый отбор по фенотипу может быть более эффективным. Примерами могут служить массовый отбор на позднее распускание елей в местах, где часто бывают поздневесенние заморозки; массовый отбор завезенных из средней Европы экологических рас елейна устойчивость к зимним морозам в Средней Швеции; массовый отбор сосны веймутовой на устойчивость к пузырчатой ржавчине.
Во всех этих случаях удавалось достичь значительного повышения устойчивости, что говорит о перспективности применения массового отбора для этих целей.
Дата добавления: 2017-08-01; просмотров: 1131;