Кодирование информации.
Кодирование – это преобразование информации в условную форму — код. Кодирование происходит двоичным кодом, т.е. наличием или отсутствием электрического импульса в тот или иной момент времени, такая форма кодирования крайне проста и устойчива к помехам. Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп или «пачек» импульсов («залпов» импульсов). Параметры этих импульсов могут быть одинаковыми, но число импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «рисунок» пачки различны и зависят от характеристик стимула. Кодирование обеспечивается также числом одновременно возбужденных нейронов, местом возбуждения в нейронном слое.
Особенности кодирования.
В отличие от телефонных или телевизионных кодов, которые декодируются восстановлением первоначального сообщения в исходном виде, в сенсорной системе такого декодирования не происходит. Особенностью нервного кодирования является множественность и перекрытие кодов. Один и тот же сигнал (например, его интенсивность) может иметь несколько кодов: частота и число импульсов в пачке, число возбужденных нейронов и их локализация в слое участков ЦНС. В коре большого мозга сигналы кодируются последовательностью включения параллельно работающих нейронных каналов, синхронностью ритмических импульсных разрядов, изменением их числа. В коре может быть позиционное кодирование, когда какой-то признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в определенном месте нейронного слоя. Например, возбуждение локальной группы нейронов зрительной коры указывает на то, что в определенной части поля зрения появился свет определенного размера и ориентации. Для периферических отделов сенсорной системы типично временное кодирование признаков раздражителя, а на высших уровнях происходит переход к преимущественно пространственному (в основном позиционному) коду.
Детектирование сигналов - избирательное выделение нейроном того или иного признака раздражителя. Такой анализ осуществляют нейроны-детекторы, они избирательно реагируют лишь на определенные параметры стимула. Например, нейрон зрительной области коры отвечает разрядом лишь на один признак, расположенной в определенной части поля зрения. При других видах воздействия ответят другие нейроны. В коре больших полушарий сконцентрированы детекторы сложных признаков и целых образов и явлений. Многие детекторы имеют генетическую природу, а часть могут развиться в онтогенезе каждого индивидуума.
Опознание образов является сложным процессом. Синтезируя сигналы от нейронов-детекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его с множеством образов, хранящихся в памяти. Опознание произойдет тем быстрее и полнее, чем полнее информация о событиях, явлениях и образах, хранящаяся в памяти. Опознание завершается формированием восприятия, и затем заключения о том, с кем или с чем встретился организм, т.е. мы осознаем, что или кого видим, какой цвет или запах действуют. Опознание часто происходит независимо от изменчивости сигнала. Мы хорошо различаем, например, предметы при различной их освещенности, окраске, размере, положении в поле зрения, т.е. сенсорная система формирует независимый от изменений ряда признаков сигнала сенсорный образ.
Переработка информации в сенсорной системе происходит в результате взаимодействия процессов возбуждения и торможения и межнейронного взаимодействия. Возбудительное взаимодействие заключается в том, что аксон каждого нейрона, контактирует с несколькими нейронами, каждый из которых получает сигналы от нескольких клеток предыдущего слоя. Совокупность рецепторов, сигналы которых поступают на данный нейрон, называют его рецептивным полем. Эти поля соседних нейронов частично перекрываются. В результате такой организации связей в сенсорной системе образуется так называемая нервная сеть. В результате чего повышается чувствительность системы к слабым сигналам и обеспечивается высокая приспособляемость к меняющимся условиям среды.
Тормозная переработка сенсорной информации основана на том, что обычно каждый возбужденный сенсорный нейрон активирует тормозный интернейрон. Интернейрон в свою очередь подавляет импульсацию как самого возбудившего его элемента (последовательное, или возвратное, торможение), так и его соседей по слою (боковое, или латеральное, торможение). Торможение выражено тем больше, чем сильнее возбужден первый элемент и чем ближе к нему соседняя клетка. Значительная часть операций по снижению избыточности и выделению наиболее существенных признаков о раздражителе производится латеральным торможением.
Адаптация органов чувств заключается в способности их приспосабливать свои свойства к условиям окружающей среды и потребностям организма. Адаптация заключается в снижении абсолютной и повышении дифференциальной чувствительности сенсорной системы. Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя (например, мы не замечаем запах при длительном действии его, снижается интенсивность воздействия яркого света – адаптация к свету или, наоборот, адаптация к темноте и т.д.). Адаптация начинается с рецепторов, а затем охватываются все нейронные уровни сенсорной системы. По скорости развития адаптации все рецепторы делятся на быстро- и медленно адаптирующиеся. Быстро адаптирующиеся рецепторы после развития адаптации практически не посылают в мозг информации о длящемся раздражении. Вторые эту информацию передают в значительно ослабленном виде. После прекращения действия раздражителя, абсолютная чувствительность (порог) восстанавливается. В сенсорной адаптации важную роль играют эфферентные влияния сенсорной системы, т.е. нисходящие влияния более высоких на низкие ее отделы. Эфферентные влияния в сенсорных системах чаще всего имеют тормозной характер, т.е. приводят к уменьшению их чувствительности и ограничивают поток афферентных сигналов. Однако этот процесс может иметь противоположный характер в том случае, если адаптация происходит с повышением чувствительности (например, адаптация к темноте).
Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровнях. Особенно широка интеграция сигналов в ретикулярной формации. В коре большого мозга происходит интеграция сигналов высшего порядка. В результате образования множественных связей с другими сенсорными и неспецифическими системами многие корковые нейроны приобретают способность отвечать на сложные комбинации сигналов разной модальности. Это особенно свойственно нервным клеткам ассоциативных областей коры больших полушарий, которые обладают высокой пластичностью, что обеспечивает перестройку их свойств в процессе непрерывного обучения опознанию новых раздражителей. Взаимодействие анализаторов идет по двум видам: 1) когда анализатор угнетает функцию другого анализатора и 2) анализатор усиливает функцию другого анализатора. Например, при нарушении функции зрения, у слепых тактильный и слуховой анализаторы имеют более высокую чувствительность, чем у людей с нормальным зрением. Взаимодействие и адаптация анализаторов происходят уже на уровне подкорковых структур, т.к. на уровне этих образований есть специализированные нейроны, реагирующие на определенные модальности конкретного раздражителя. Подкорковые структуры избирательно пропускают те или иные импульсы с периферии. Они блокируют («фильтруют») множество импульсов, способствуя тем самым вычленить из массы раздражителей наиболее важные в биологическом отношении (биологическая модальность) или наиболее сильные (сенсорная модальность) воздействия. Лишь после этого дается оценка происходящим явлениям и формируется адекватная поведенческая реакция с вовлечением соматических и вегетативных компонентов, т.е. формируется по ходу реакции функциональная система. Она направлена на достижение полезного приспособительного результата. Импульсы с периферии идут по двум каналам: 1)специфический путь, свойственный для данного органа чувств. Этот путь несет импульсы к соответствующему корковому представительству (первичная проекционная зона), 2) неспецифический путь несет импульсы с участием ретикулярной формации ствола мозга. Благодаря взаимному влиянию нейрональных сетей разных уровней ЦНС происходит взаимодействие разных органов чувств. Все это способствует формированию адекватного поведения.
ЗРЕНИЕ.
Глазное яблоко имеет шарообразную форму. Глаз имеет несколько оболочек: наружная, непрозрачная соединительнотканная оболочка – склера, в передней части глаза она переходит в прозрачную роговую оболочку. Под склерой лежит сосудистая оболочка. Спереди эта оболочка утолщается и переходит в начале в ресничное тело и далее в радужную оболочку. В центре радужной оболочки имеется отверстие – зрачок. С внутренней стороны сосудистой оболочки находится слой клеток пигментного эпителия, а к нему прилегает самая внутренняя оболочка – сетчатая оболочка, или ретина. Ретина выполняет самую основную функцию глаза – преобразование светового раздражения в нервное возбуждение и первичную обработку сигнала. Между роговой и радужной оболочками имеется полость, наполненная жидкостью – передняя камера глаза. За радужной оболочкой находится прозрачное тело в виде двояковыпуклой линзы – хрусталик. Он прикрепляется к ресничному телу. За хрусталиком вся полость заполнена стекловидным телом. Первичная обработка информации происходит в сетчатке, которая состоит из 10 слоев. Первые 4 слоя образованы фоторецепторами. Нейроны сетчатки – горизонтальные, биполярные, амакриновые и ганглиозные клетки – неоднородны по величине и форме, толщине, длине, количеству отростков и синаптических образований. На пути к сетчатке лучи света проходят через несколько прозрачных сред — роговицу, хрусталик и стекловидное тело. Они входят в понятие – преломляющая или оптическая система глаза. Оптическая сила глаза измеряется в диоптриях. Преломляющая сила здорового глаза составляет 59D при рассматривании далеких и 70,5 D — при рассматривании близких предметов.
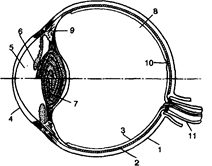
Механизм аккомодации (по Гельмгольцу): 1 – склера, 2 – сосудистая оболочка, 3 – сетчатка, 4 – роговица,
5 – передняя камера, 6 – радужная оболочка, 7 – хрусталик, 8 - стекловидное тело; 9 — ресничная мышца, ресничные отростки и ресничный поясок (цинновы связки); 10 — центральная ямка; 11 — зрительный нерв.
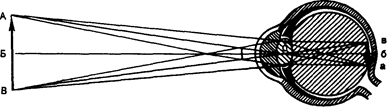
Рис. 14.3. Ход лучей от объекта и построение изображения на сетчатой оболочке глаза.
АВ — предмет; ав — его изображение; 0 — узловая точка; Б-б — главная оптическая ось
У здоровых людей преломляющая сила глаза колеблется в пределах 52–64 D. Наибольшую преломляющую силу имеет роговица (около 43 D) и несколько меньше хрусталик (около19 D). На сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево. Если смотреть вдаль и одновременно представить себе насколько ясно виден предмет, находящийся на расстоянии 20–30 см перед глазом, то легко заметить, что он виден не резко и расплывчато. Причина этого явления состоит в том, что, когда на сетчатке находится изображение далеко расположенного предмета, то лучи света от ближнего предмета сходятся к точкам, лежащим за сетчатой. Возникает так называемый круг светорассеяния и нечеткое изображение предмета.
Аккомодация.
Аккомодацией называют приспособление глаза к ясному видению объектов, расположенных на различном расстоянии. Когда мы смотрим на далекие предметы их изображение сфокусировано на сетчатке и они видны ясно, а изображение близких предметов при этом расплывчато, так как лучи от них собираются за сетчаткой. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну, следовательно, преломляющую способность. При рассматривании близких предметов хрусталик делается более выпуклым и лучи, расходящиеся от какой-либо точки объекта, сходятся на сетчатке. Аккомодация глаз происходит за счет сокращения ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, которую всегда растягивают, т.е. уплощают, волокна ресничного пояска (циннова связка). Сокращение гладких мышечных клеток ресничного тела уменьшает тягу цинновых связок, что увеличивает выпуклость хрусталика в силу его эластичности. Пределы возможностей аккомодации (ширина) определяется тем, на каких расстояниях от глаза находятся точки, резко видимые при полностью расслабленной аккомодации (дальняя точка ясного видения), и точка видимая резко при максимальном напряжении аккомодации глаз (ближняя точка ясного видения). После достижения ~ 10 лет дальняя точка довольно стабильна, а ближняя с годами отодвигается все дальше, а максимальная сила аккомодации постепенно уменьшается. Если в 10 лет сила аккомодации составляет примерно 14±2,5 D, то к 50 годам – 2±1 и в 60 лет – всего 1±0,5. В указанные пределы укладываются более 95% здоровых глаз. Для ясного видения объекта необходимо, чтобы он был сфокусирован на сетчатке. В пожилом возрасте аккомодационная способность снижается, и это явление называется старческой дальнозоркостью или пресбиопией. Пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами. Причиной таких изменений является уменьшение эластичности хрусталика. Для нормального глаза молодого человека дальняя точка ясного видения лежит в бесконечности. Далекие предметы он рассматривает без всякого напряжения аккомодации, т.е. без сокращения ресничной мышцы. Ближайшая точка ясного видения находится на расстоянии 10 см от глаза. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, ограничивает аккомодацию глаза при рассматривании близких предметов. Наоборот, парасимпатомиметические
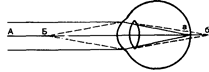
Ход лучей от близкой и далекой точек
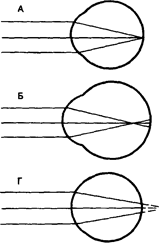
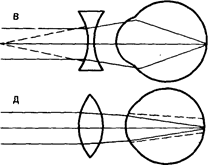
Рефракция в нормальном (А), близоруком (Б) и дальнозорком (Г) глазу и оптическая коррекция близорукости (В) и дальнозоркости (Д) (схема).
Дата добавления: 2015-12-11; просмотров: 1017;