Органоиды клетки, их строение и функции.
Органоиды – постоянные и обязательные компоненты клеток; специализированные участки цитоплазмы клетки, имеющие определенную структуру и выполняющие определенные функции в клетке. Различают органоиды общего и специального назначения.
Органоиды общего назначения имеются в большинстве клеток (эндоплазматическая сеть, митохондрии, пластиды, комплекс Гольджи, лизосомы, вакуоли, клеточный центр, рибосомы). Органоиды специального назначения характерны только для специализированных клеток (миофибриллы, жгутики, реснички, сократительные и пищеварительные вакуоли). Органоиды (за исключением рибосом и клеточного центра) имеют мембранное строение.
Эндоплазматическая ретикулум(ЭПР) – это разветвленная система соединенных между собой полостей, трубочек и каналов, образованных элементарными мембранами и пронизывающая всю толщу клетки. Открыта в 1943 г. Портером. Особенно много каналов эндоплазматической сети в клетках с интенсивным обменом веществ. В среднем объем ЭПС составляет от 30% до 50% общего объема клетки. ЭПС лабильна. Форма внутренних лакун и кана
лов, их размер, расположение в клетке и количество изменяются в процессе жизнедеятельности. Развита сильнее в животных клетка. ЭПС морфологически и функционально связана с пограничным слоем цитоплазмы, ядерной оболочкой, рибосомами, комплексом Гольджи, вакуолями, образуя вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и передвижения веществ внутри клетки. Вблизи эндоплазматической сети накапливаются митохондрии и пластиды.
Выделяют две разновидности ЭПС: шероховатую и гладкую. На мембранах гладкой (агранулярной) ЭПС локализованы ферменты систем жирового и углеводного синтеза: здесь происходит синтез углеводов и почти всех клеточных липидов. Мембраны гладкой разновидности эндоплазматической сети преобладают в клетках сальных желез, печени (синтез гликогена), в клетках с большим содержанием питательных веществ (семена растений). На мембране шероховатой (гранулярной) ЭПС располагаются рибосомы, где осуществляется биосинтез белков. Часть синтезируемых ими белков включается в состав мембраны эндоплазматической сети, остальные поступают в просвет ее каналов, где преобразуются и транспортируются в комплекс Гольджи. Особенно много шероховатых мембран в клетках желез и нервных клетках.
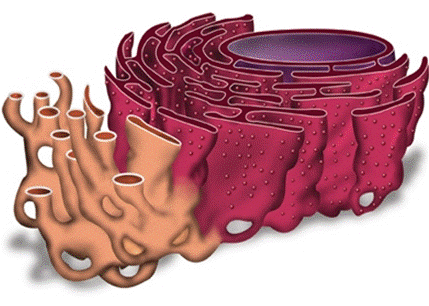
Рис. Шероховатый и гладкий эндоплазматический ретикулум.
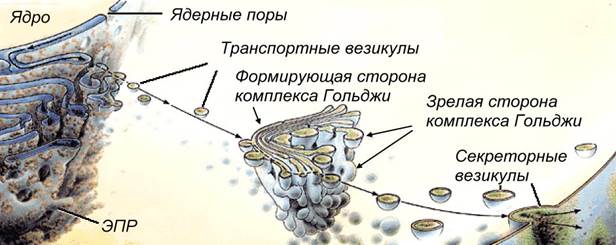
Рис. Транспорт веществ по системе ядро – эндоплазматический ретикулум (ЭПР) – комплекс Гольджи.
Функции эндоплазматической сети:
1) синтез белков (шероховатая ЭПС), углеводов и липидов (гладкая ЭПС);
2) транспорт веществ, как поступивших в клетку, так и вновь синтезируемых;
3) деление цитоплазмы на отсеки (компартменты), что обеспечивает пространственное разделение ферментных систем, необходимое для их последовательного вступления в биохимические реакции.
Митохондрии – присутствуют практически во всех типах клеток одно- и многоклеточных организмов (за исключением эритроцитов млекопитающих). Число их в разных клетках варьирует и зависит от уровня функциональной активности клетки. В клетке печени крысы их около 2500, а в мужской половой клетке некоторых моллюсков – 20 – 22. Их больше в грудной мышце летающих птиц, чем в грудной мышце нелетающих.
Митохондрии имеют форму сферических, овальных и цилиндрических телец. Размеры составляют 0,2 – 1,0 мкм и диаметре и до 5 - 7 мкм в длину.
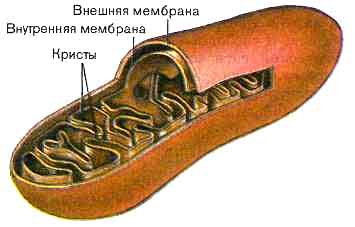
Рис. Митохондрия.
Длина нитевидных форм достигает 15-20 мкм. Снаружи митохондрии ограничены гладкой наружной мембраной, сходной по составу с плазмалеммой. Внутренняя мембрана образует многочисленные выросты – кристы – и содержит многочисленные ферменты, АТФ-сомы (грибовидные тела), участвующие в процессах трансформации энергии питательных веществ в энергию АТФ. Количество крист зависит от функции клетки. В митохондриях мышц крист очень много, они занимают всю внутреннюю полость органоида. В митохондриях эмбриональных клеток кристы единичны. В растительных выросты внутренней мембраны чаще имеют форму трубочек. Полость митохондрии заполнена матриксом, в котором содержатся вода, минеральные соли, белки-ферменты, аминокислоты. Митохондрии имеют автономную белоксинтезирующую систему: кольцевую молекулу ДНК, различные виды РНК и более мелкие, чем в цитоплазме рибосомы.
Митохондрии тесно связаны мембранами эндоплазматической сети, каналы которой часто открываются прямо в митохондрии. При повышении нагрузки на орган и усилении синтетических процессов, требующих затраты энергии, контакты между ЭПС и митохондриями становятся особенно многочисленными. Число митохондрий может быстро увеличиваться путем деления. Способность митохондрий к размножению обусловлена присутствием в них молекулы ДНК, напоминающей кольцевую хромосому бактерий.
Функции митохондрий:
1) синтез универсального источника энергии – АТФ;
2) синтез стероидных гормонов;
3) биосинтез специфических белков.
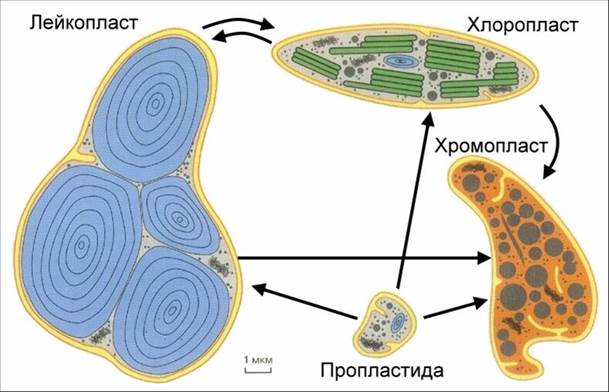
Пластиды – органоиды мембранного строения, характерные только для растительных клеток. В них происходят процессы синтеза углеводов, белков и жиров. По содержанию пигментов их делят на три группы: хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют относительно постоянную эллиптическую или линзовидную форму. Размер по наибольшему диаметру составляет 4 – 10 мкм. Количество в клетке колеблется от нескольких единиц до нескольких десятков. Их размер, интенсивность окраски, количество и расположение в клетке зависят от условий освещения, вида и физиологического состояния растений.
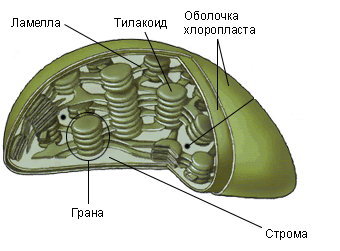
Рис. Хлоропласт, строение.
Это белково-липоидные тела, состоящие на 35-55% из белка, 20-30% - липидов, 9% - хлорофилла, 4-5% каратиноидов, 2-4% нуклеиновых кислот. Количество углеводов варьирует; обнаружено некоторое количество минеральных веществ Хлорофилл – сложный эфир органической двухосновной кислоты – хлорофиллина и органических спиртов – метилового (СН3ОН) и фитола (С20Н39ОН). У высших растений в хлоропластах постоянно присутствуют хлорофилл а – имеет сине-зеленую окраску, и хлорофилл b – желто-зеленую; причем содержание хлорофилла, а в несколько раз больше.
Кроме хлорофилла в состав хлоропластов входят пигменты - каротин С40Н56 и ксантофилл С40Н56О2 и некоторые другие пигменты (каратиноиды). В зеленом листе желтые спутники хлорофилла маскируются более яркой зеленой окраской. Однако осенью, при листопаде, у большинства растений хлорофилл разрушается и тогда обнаруживается присутствие в листе каратиноида – лист становится желтым.
Хлоропласт одет двойной оболочкой, состоящей из наружной и внутренней мембран. Внутреннее содержимое – строма – имеет ламеллярное (пластинчатое) строение. В бесцветной строме выделяют граны – окрашенные в зеленые цвет тельца, 0,3 – 1,7 мкм. Они представляют собой совокупность тилакоидов – замкнутых телец в виде плоских пузырьков или дисков мембранного происхождения. Хлорофилл в виде мономолекулярного слоя располагается между белковым и липидным слоями в тесной связи с ними. Пространственное расположение молекул пигментов в мембранных структурах хлоропластов является весьма целесообразным и создает оптимальные условия для наиболее эффективного поглощения, передачи и использования лучистой энергии. Липиды образуют безводные диэлектрические слои мембран хлоропласта, необходимые для функционирования электронно-транспортной цепи. Роль звеньев цепи переноса электронов выполняют белки (цитохромы, пластохиноны, ферредоксин, пластоцианин) и отдельные химические элементы – железо, марганец и др. Количество гран в хлоропласте от 20 до 200. Между гранами, связывая их друг с другом, располагаются ламеллы стромы. Ламеллы гран и ламеллы стромы имеют мембранное строение.
Внутренне строение хлоропласта делает возможным пространственное разобщение многочисленных и разнообразных реакций, составляющих в своей совокупности содержание фотосинтеза.
Хлоропласты, как и митохондрии, содержат специфическую РНК и ДНК, а также более мелкие рибосомы и весь молекулярный арсенал, необходимый для биосинтеза белка. У этих органоидов имеется достаточное для обеспечения максимальной активности белоксинтезирующей системы количество и-РНК. Вместе с тем в них содержится и достаточно ДНК для кодирования определенных белков. Они размножаются делением, путем простой перетяжки.
Установлено, что хлоропласты могут изменять свою форму, размеры и положение в клетке, т. е. способны самостоятельно двигаться (таксис хлоропластов). В них обнаружено два типа сократительных белков, за счет которых, очевидно, и осуществляется активное движение этих органоидов в цитоплазме.
Хромопласты широко распространены в генеративных органах растений. Они окрашивают лепестки цветков (лютика, георгина, подсолнечника), плоды (томатов, рябины, шиповника) в желтый, оранжевый, красный цвета. В вегетативных органах хромопласты встречаются значительно реже.
Окраска хромопластов обусловлена присутствием каратиноидов – каротина, ксантофилла и ликопина, которые в пластидах находятся в различном состоянии: в виде кристаллов, липоидного раствора или в соединении с белками.
Хромопласты, по сравнению с хлоропластами, имеют более простое строение – в них отсутствует ламеллярная структура. Химический состав также отличен: пигменты – 20–50%, липиды до 50%, белки – около 20%, РНК – 2-3%. Это свидетельствует о меньшей физиологической активности хлоропластов.
Лейкопласты не содержат пигментов, они бесцветны. Эти самые мелкие пластиды имеют округлую, яйцевидную или палочковидную форму. В клетке они часто группируются вокруг ядра.
Внутренне структура, еще менее дифференцирована по сравнению с хлоропластами. В них осуществляется синтез крахмала, жиров, белков. В соответствии с этим выделяют три вида лейкопластов – амилопласты (крахмал), олеопласты (растительные масла) и протеопласты (белки).
Возникают лейкопласты из пропластид, с которыми они сходны по форме и строению, а отличаются лишь размерами.
Все пластиды генетически связаны друг с другом. Они образуются из пропластид – мельчайших бесцветных цитоплазматических образований, сходных по внешнему виду с митохондриями. Пропластиды находятся в спорах, яйцеклетках, в эмбриональных клетках точек роста. Непосредственно из пропластид образуются хлоропласты (на свету) и лейкопласты (в темноте), а из них развиваются хромопласты, являющиеся конечным продуктом в эволюции пластид в клетке.
Комплекс Гольджи – впервые был обнаружен в 1898 г. Итальянским ученым Гольджи в животных клетках. Это система внутренних полостей, цистерн (5-20), располагающихся сближено и параллельно друг другу, и крупных и мелких вакуолей. Все эти образования имеют мембранное строение и являются специализированными участками эндоплазматической сети. В животных клетках комплекс Гольджи развит лучше, чем в растительных; в последних он называется диктиосомы.
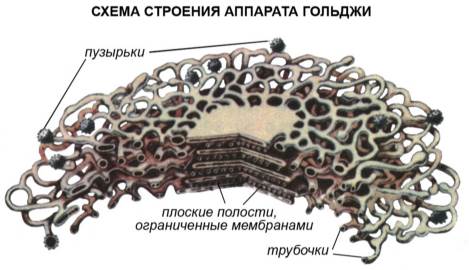
Рис. Строение комплекса Гольджи.
Попадающие в пластинчатый комплекс белки и липиды, подвергаются различным преобразованиям, накапливаются, сортируются, упаковываются в секреторные пузырьки и транспортируются по назначению: к различным структурам внутри клетки или за пределы клетки. Мембраны комплекса Гольджи также синтезируют полисахариды и образуют лизосомы. В клетках молочных желез комплекс Гольджи участвует в образовании молока, а в клетках печени – желчи.
Функции комплекса Гольджи:
1) концентрация, обезвоживание и уплотнение синтезированных в клетке белков, жиров, полисахаридов и веществ, поступивших извне;
2) сборка сложных комплексов органических веществ и подготовка их к выведению из клетки (целлюлоза и гемицеллюлоза у растений, гликопротеины и гликолипиды у животных);
3) синтез полисахаридов;
4) образование первичных лизосом.
Лизосомы - небольшие овальные тельца диаметром 0,2-2,0 мкм. Центральное положение занимает вакуоль, содержащая 40 (по разным данным 30-60) гидролитических ферментов, способных в кислой среде (рН 4,5-5) расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и другие вещества.
Вокруг этой полости располагается строма, одетая снаружи элементарной мембраной. Расщепление веществ при помощи ферментов называется лизисом, поэтому органоид назван лизосомой. Образование лизосом происходит в комплексе Гольджи. Первичные лизосомы приближаются непосредственно к пиноцитозным или фагоцитозным вакуолям (эндосомам) и изливают свое содержимое в их полость, образуя вторичные лизосомы (фагосомы), внутри которых и происходит переваривание веществ. Продукты лизиса через мембрану лизосом поступают в цитоплазму и включаются в дальнейший обмен веществ. Вторичные лизосомы с остатками непереваренных веществ называются остаточными тельцами. Примером вторичных лизосом являются пищеварительные вакуоли простейших.
Функции лизосом:
1) внутриклеточное переваривание макромолекул пищи и чужеродных компонентов, поступающих в клетку при пино- и фагоцитозе, обеспечивая клетку дополнительным сырьем для биохимических и энергетических процессов;
2) при голодании лизосомы переваривают некоторые органоиды и на какое-то время пополняют запас питательных веществ;
3) разрушение временных органов эмбрионов и личинок (хвост и жабры у лягушки) в процессе постэмбрионального развития;
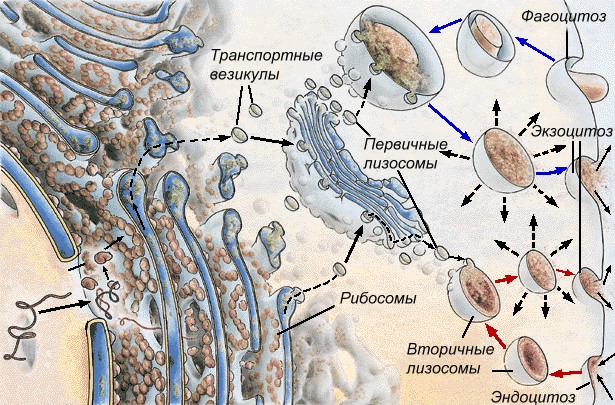
Рис. Образование лизосом
Вакуоли – полости в цитоплазме растительных клеток и протист, заполненные жидкостью. Имеют форму пузырьков, тонких канальцев и другую. Вакуоли образуются из расширений эндоплазматической сети и пузырьков комплекса Гольджи как тончайшие полости, затем по мере роста клетки и накопления продуктов обмена объем их увеличивается, а количество сокращается. Развитая сформировавшаяся клетка имеет обычно одну большую вакуоль, занимающую центральное положение.
Вакуоли растительных клеток заполнены клеточным соком, который представляет собой водный раствор органических (яблочная, щавелевая, лимонная кислоты, сахара, инулин, аминокислоты, белки, дубильные вещества, алкалоиды, глюкозиды) и минеральных (нитраты, хлориды, фосфаты) веществ.
У протист встречаются пищеварительные вакуоли и сократительные.
Функции вакуолей:
1) хранилища запасных питательных веществ и вместилища выделений ( у растений);
2) определяют и поддерживают осмотическое давление в клетках;
3) обеспечивают внутриклеточное пищеварение у протист.
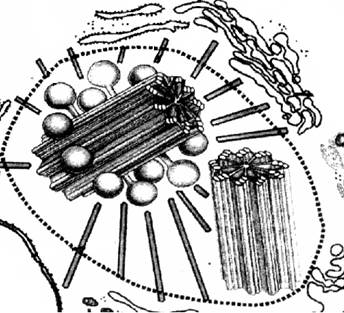
Рис. Клеточный центр.
Клеточный центр обычно находится вблизи ядра и состоит из двух центриолей, расположенных перпендикулярно друг другу и окруженных лучистой сферой. Каждая центриоль представляет собой полое цилиндрическое тельце длиной 0,3-0,5 мкм и длиной 0,15 мкм, стенка которого образована 9 триплетами микротрубочек. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем.
Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу. Они формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками, являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам. В клетках высших растений клеточный центр центриолей не имеет.
Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли.
Центросома характерна для клеток животных, некоторых грибов, водорослей, мхов и папоротников.
Функции клеточного центра:
1) образование полюсов деления и формирование микротрубочек веретена деления.
Рибосомы - мелкие сферические органоиды, от 15 до 35 нм. Состоят из двух субъединиц большой (60S) и малой (40S). Содержат около 60% белка и 40% рибосомальной РНК. Молекулы рРНК образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка. Субъединицы рибосом образуются в ядрышках. и через поры в ядерной оболочке поступают в цитоплазму, где располагаются либо на мембране ЭПА, либо на наружной стороне ядерной оболочки, либо свободно в цитоплазме. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка. По сравнению с митохондриями, пластидами, клетками прокариот рибосомы в цитоплазме эукариотических клеток крупнее. Могут объединяться по 5-70 единиц в полисомы.
Функции рибосом:
1) участие в биосинтезе белка.
Рис. 287. Рибосома: 1 — малая субъединица; 2 — большая субъединица.
Реснички, жгутики – выросты цитоплазмы, покрытые элементарной мембраной, под которой находится 20 микротрубочек, образующих 9 пар по периферии и две одиночные в центре. У основания ресничек и жгутиков расположены базальные тельца. Длина жгутиков достигает 100 мкм. Реснички – это короткие – 10-20 мкм – жгутики. Движение жгутиков винтовое, а ресничек – веслообразное. Благодаря ресничкам и жгутикам передвигаются бактерии, протисты, ресничные, перемещаются частицы или жидкости (реснички мерцательного эпителия дыхательных путей, яйцеводов), половые клетки (сперматозоиды).
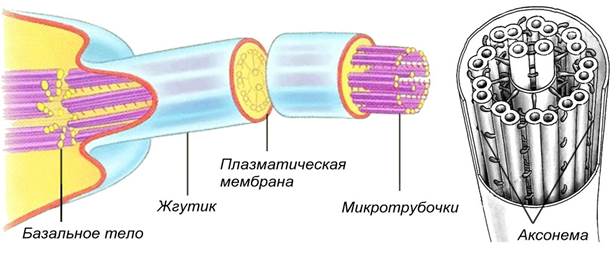
Рис. Строение жгутиков и ресничек эукариот
Включения — временные компоненты цитоплазмы, то возникающие, то исчезающие. Как правило, они содержатся в клетках на определенных этапах жизненного цикла. Специфика включений зависит от специфики соответствующих клеток тканей и органов. Включения встречаются преимущественно в растительных клетках. Они могут возникать в гиалоплазме, различных органеллах, реже в клеточной стенке.
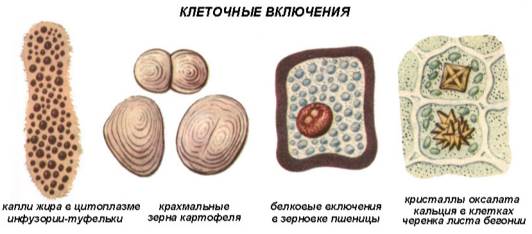
В функциональном отношении включения представляют собой либо временно выведенные из обмена веществ клетки соединения (запасные вещества — крахмальные зерна, липидные капли и отложения белков), либо конечные продукты обмена (кристаллы некоторых веществ).
Крахмальные зерна. Это наиболее распространенные включения растительных клеток. Крахмал запасается у растений исключительно в виде крахмальных зерен. Они образуются только в строме пластид живых клеток. В процессе фотосинтеза в зеленых листьях образуется ассимиляционный, или первичный крахмал. Ассимиляционный крахмал в листьях не накапливается и, быстро гидролизуясь до сахаров, оттекает в части растения, в которых происходит его накопление. Там он вновь превращается в крахмал, который называют вторичным. Вторичный крахмал образуется и непосредственно в клубнях, корневищах, семенах, то есть там, где он откладывается в запас. Тогда его называют запасным. Лейкопласты, накапливающие крахмал, называют амилопластами. Особенно богаты крахмалом семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений.
Липидные капли. Встречаются практически во всех растительных клетках. Наиболее богаты ими семена и плоды. Жирные масла в виде липидных капель — вторая по значению (после крахмала) форма запасных питательных веществ. Семена некоторых растений (подсолнечник, хлопчатник и т.д.) могут накапливать до 40% масла от массы сухого вещества.
Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. Липидные капли могут накапливаться и в лейкопластах, которые называют элайопластами.
Белковые включения образуются в различных органеллах клетки в виде аморфных или кристаллических отложений разнообразной формы и строения. Наиболее часто кристаллы можно встретить в ядре — в нуклеоплазме, иногда в перинуклеарном пространстве, реже в гиалоплазме, строме пластид, в расширениях цистерн ЭПР, матриксе пероксисом и митохондриях. В вакуолях встречаются как кристаллические, так и аморфные белковые включения. В наибольшем количестве кристаллы белка встречаются в запасающих клетках сухих семян в виде так называемых алейроновых3 зерен или белковых телец.
Запасные белки синтезируются рибосомами во время развития семени и откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, и белок кристаллизуется. В результате этого в зрелом сухом семени белковые вакуоли превращаются в белковые тельца (алейроновые зерна).
Включения классифицируют на трофические, представляющие собой запасы питательных веществ (углеводные – гликоген в клетках печени и мышцах, белковые – желточные включения в яйцеклетках, жировые – капли жира в клетках подкожной жировой клетчатки), секреторные продукты жизнедеятельности клеток желез внешней и внутренней секреции (ферменты, гормоны, слизь и др.), пигментные – содержат различные пигменты (меланин, липофусцин, гемоглобин и др.) и экскреторные – продукты обмена веществ, подлежащие удалению из клетки, (кристаллы щавелевой кислоты, щавелевокислого кальция, мочевина)
Дата добавления: 2015-11-04; просмотров: 11910;