Клеточные мембраны
В основе структурной организации клетки лежит мембранный принцип строения, то есть клетка в основном построена из мембран. Все биологические мембраны имеют общие структурные особенности и свойства.
В настоящее время общепринята жидкостно-мозаичная модель строения мембраны.
| Химический состав и строение мембраны |
Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Липиды составляют в среднем ≈40% химического состава мембраны. В бислое хвосты молекул в мембране обращены друг к другу, а полярные головки — наружу, поэтому поверхность мембраны гидрофильна. Липиды определяют основные свойства мембран.
Помимо липидов в состав мембраны входят белки (в среднем ≈60%). Они определяют большинство специфических функций мембраны. Молекулы белков не образуют сплошного слоя (рис. 280). В зависимости от локализации в мембране различают:
© периферические белки — белки, располагающиеся на наружной или внутренней поверхности липидного бислоя;
© полуинтегральные белки — белки, погруженные в липидный бислой на различную глубину;
© интегральные, или трансмембранные белки — белки, пронизывающие мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки.
Мембранные белки могут выполнять различные функции:
© транспорт определенных молекул;
© катализ реакций, происходящих на мембранах;
© поддержание структуры мембран;
© получение и преобразование сигналов из окружающей среды.
|
 Рис. 280. Цитоплазматическая мембрана:
1 — липидный бислой; 2 — погруженные белки; 3 — периферические белки; 4 — интегральные белки; 5 — олигосахаридные цепи.
Рис. 280. Цитоплазматическая мембрана:
1 — липидный бислой; 2 — погруженные белки; 3 — периферические белки; 4 — интегральные белки; 5 — олигосахаридные цепи.
В состав мембраны может входить от 2 до 10% углеводов. Углеводный компонент мембран обычно представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Функции углеводов клеточной мембраны до конца не выяснены, однако можно сказать, что они обеспечивают рецепторные функции мембраны.
В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину в несколько десятков нанометров. В нем происходит внеклеточное пищеварение, располагаются многие рецепторы клетки, с его помощью, по-видимому, происходит адгезия клеток.
Молекулы белков и липидов подвижны, способны перемещаться, главным образом, в плоскости мембраны. Мембраны асимметричны, то есть липидный и белковый состав наружной и внутренней поверхности мембраны различен.
Толщина плазматической мембраны в среднем 7,5 нм.
| Функции мембран |
Клеточные мембраны играют важную роль по ряду причин:
© они отделяют клеточное содержимое от внешней среды;
© регулируют обмен между клеткой и средой;
© делят клетки на отсеки, или компартменты, предназначенные для тех или иных специализированных метаболических путей;
© некоторые химические реакции протекают на самих мембранах (световые реакции фотосинтеза в хлоропластах, окислительное фосфорилирование при дыхании в митохондриях);
© обеспечивают связь между клетками в тканях многоклеточных организмов;
© на мембранах располагаются рецепторные участки для распознавания внешних стимулов.
| Транспорт веществ через мембрану |
Одна из основных функций мембраны — транспортная, обеспечивающая обмен веществ между клеткой и внешней средой. Мембраны обладают свойством избирательной проницаемости, то есть хорошо проницаемы для одних вещества или молекул и плохо проницаемы (или совсем непроницаемы) для других. Проницаемость мембран для разных веществ зависит и от свойств их молекул (полярность, размер и т.д.), и от характеристики мембран (внутренняя часть липидного слоя гидрофобна).
Существуют различные механизмы транспорта веществ через мембрану (рис. 281). В зависимости от необходимости использования энергии для осуществления транспорта веществ, различают:
© пассивный транспорт — транспорт веществ, идущий без затрат энергии;
© активный транспорт — транспорт, идущий с затратами энергии.
| Пассивный транспорт |
В основе пассивного транспорта лежит разность концентраций и зарядов. При пассивном транспорте вещества всегда перемещаются из области с более высокой концентрацией в область с более низкой, то есть по градиенту концентрации. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Различают три основных механизма пассивного транспорта:
© Простая диффузия — транспорт веществ непосредственно через липидный бислой. Через него легко проходят газы, неполярные или малые незаряженные полярные молекулы. Чем меньше молекула и чем более она жирорастворима, тем быстрее она проникает через мембрану. Интересно, что вода, несмотря на то, что она относительно нерастворима в жирах, очень быстро проникает через липидный бислой. Это объясняется тем, что ее молекула мала и электрически нейтральна. Диффузию воды через мембраны называют осмосом.
©
|
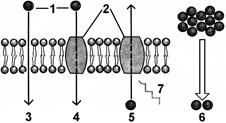 Рис. 281. Транспорт веществ через мембрану:
1 — транспортируемая молекула; 2 — транспортный белок; 3 — диффузия; 4 — облегченная диффузия; 5 — активный транспорт; 6 — электрохимический градиент; 7 — энергия.
Рис. 281. Транспорт веществ через мембрану:
1 — транспортируемая молекула; 2 — транспортный белок; 3 — диффузия; 4 — облегченная диффузия; 5 — активный транспорт; 6 — электрохимический градиент; 7 — энергия.
© Облегченная диффузия — транспорт веществ с помощью специальных
транспортных белков, каждый из которых отвечает за транспорт определенных молекул или групп родственных молекул. Они взаимодействуют с молекулой переносимого вещества и каким-либо способом перемещают ее сквозь мембрану. Таким образом в клетку транспортируются сахара, аминокислоты, нуклеотиды и многие другие полярные молекулы.
| Активный транспорт |
Необходимость активного транспорта возникает тогда, когда требуется обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ.
Одной из наиболее изученных систем активного транспорта является натрий-калиевый насос. Концентрация K внутри клетки значительно выше, чем за ее пределами, а Na — наоборот. Поэтому К через водяные поры мембраны пассивно диффундирует из клетки, а Na — в клетку. Вместе с тем, для нормального функционирования клетке важно поддерживать определенное соотношение ионов К и Na в цитоплазме и во внешней среде. Это оказывается возможным потому, что мембрана, благодаря наличию (Na + K)-насоса, активно перекачивает Na из клетки, а K в клетку. На работу (Na + K)-насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки.
|
 Рис. 282. Натрий-калиевый насос:
1 — присоединение ионов натрия; 2 — фосфорилирование транспортного белка; 3 — высвобождение ионов натрия; 4 — присоединение ионов калия; 5 — дефосфорилирование транспортного белка; 6 — высвобождение ионов калия.
Рис. 282. Натрий-калиевый насос:
1 — присоединение ионов натрия; 2 — фосфорилирование транспортного белка; 3 — высвобождение ионов натрия; 4 — присоединение ионов калия; 5 — дефосфорилирование транспортного белка; 6 — высвобождение ионов калия.
Насос представляет собой особый трансмембранный белок мембраны, способный к конформационным изменениям, благодаря чему он может присоединять к себе как ионы К, так и ионы Na. Цикл работы (Na + K)-насоса складывается из нескольких фаз (рис. 282):
© с внутренней стороны мембраны к белку-насосу поступают ионы Na и молекула АТФ, а с наружной — ионы K;
© ионы Na соединяются с молекулой белка, и белок приобретает АТФ-азную активность, то есть приобретает способность вызывать гидролиз АТФ, сопровождающийся освобождением энергии, приводящей в движение насос;
© освободившийся при гидролизе АТФ фосфат присоединяется к белку, то есть происходит фосфорилирование белка;
© фосфорилирование вызывает конформационные изменения белка, он оказывается неспособным удерживать ионы Na — они высвобождаются и выходят за пределы клетки;
© новая конформация белка такова, что оказывается возможным присоединение к нему ионов K;
© присоединение ионов K вызывает дефосфорилирование белка, в результате чего он вновь изменяет свою конформацию;
© изменение конформации белка приводит к высвобождению ионов K внутри клетки;
© теперь белок вновь готов присоединить к себе ионы Na.
За один цикл работы насос выкачивает из клетки 3 иона Na и закачивает 2 иона К. Такая разница в количестве переносимых ионов связана с тем, что проницаемость мембраны для ионов К выше, чем для ионов Na. Соответственно K быстрее пассивно диффундирует из клетки, чем Na в клетку.
| Эндоцитоз и экзоцитоз |
Клетка имеет механизмы, благодаря которым может осуществлять транспорт через мембрану крупных частиц и макромолекул (рис. 283). Процесс поглощения макромолекул клеткой называется эндоцитозом. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму везикул — мешочкоподобных структур, отграниченных от цитоплазмы одиночной мембраной, являющейся частью наружной цитоплазматической мембраны. Различают два типа эндоцитоза:
© фагоцитоз[13] — захват и поглощение
|
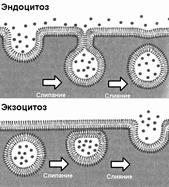 Рис 283. Эндоцитоз и экзоцитоз.
Рис 283. Эндоцитоз и экзоцитоз.
© пиноцитоз[14] — процесс захвата и поглощения капелек жидкости с растворенными в ней веществами.
Экзоцитоз — процесс выведения различных веществ из клетки. При экзоцитозе мембрана везикулы (или вакуоли), при соприкосновении с наружной цитоплазматической мембраной, сливается с ней. Содержимое везикулы выводится за пределы летки, а ее мембрана включается в состав наружной цитоплазматической мембраны.
Дата добавления: 2015-07-22; просмотров: 1998;