Рефлекторная дуга вегетативной
| Клиническая форма | Маркери інфекції | ||||
| HbsAg | HBeAg | anti-HBc | anti-HBe | anti-HBs | |
| Гострий ГВ. Розпал хвороби | + | + | IgM | - | - |
| Гострий ГВ. Реконвалесценція. | - | - | IgG, IgM | - | + |
| Хронічний ГВ. Активний проце | + | + | IgG | - | -(+) |
| Хронічний ГВ. Ремйсія. | + | - | IgG | + | -(+) |
| «Коровське окно» при гострому ГВ. | - | ± | IgМ | ± | - |
| Хронічне носійство з низькою реплікацією вірусу | -, ±, + | - | IgG | ± | - |
| Поствакцинальна реакція | - | - | - | - | + |
На наявність "свіжої" інфекції вказиваютб високі титри НВsAg і IgM до НВsAg і НВcAg. У хворих з гепатитом, що клінічно проявляється, титр НВsAg спочатку росте, а потім (у міру розвитку імунних реакцій) знижується. Антитіла до НВsAg можна виявить тільки через нескоько тижнів що пояснюється їх активним зв'язуванням в імунні комплекси. Впродовж цього періоду, т.з. "вікна" можна виявити лише антитіла до НВсAg.
Виявлення НВеAg у пацієнтів з явними ознаками хронічного гепатиту вказує на активацію процесу і подібні особи представляють високу епідемічну небезпеку.
Тестові питання
Роком народження врусології вважається
1865 рік
1892 рік
1948 РІК
Д.І. Івановський довів, що
Віруси мають один тип нуклеїнової кислоти
Віруси здатні до розмноження половим шляхом
Віруси мають розміри значно менші за розміри бактерій та здатні до розмноження виключно у тканинах організму
Вірусна частка, на її кінцевій стадіі утворення, має назву
віріон
вірус
віроїд
вірусоїд
Оболонка вірусу, що побудована з ідентичних субодиниць, має назву
капсид
нуклеокапсид
суперкапсид
мембрана
Терміном “віріон” позначається
Нуклеїнова кислота віруса в інтегрованому з геномом клітини стані
Заключна форма розвитку вірусу, яку він має виходячи із клітини
Зовніщня оболонка вірусу з антигенними комплексами
Віроїди – це
Дуже маленькі білкові молекули, що входять ло складу оболонок складних вірусів
Збудники особливо небезпечних хвороб нільських крокодилів
Одноланцюгові, кільцева РНК, не здаатна до трансляції, є збудником багатьох хвороб рослин
Пріони є збудниками
гострих респіраторних захворювань корів
губчастих енцефалопатій людини та тварин
хвороб кишково-щлункового тракту людини
Пріони інактивуються
Ультрафіолетовим опроміненням на протязі 15 хвилин
хлораміном на протязв 5 хвилин
Автоклавуванням при 136 градусах на протязі 18 хвилин /
2% розчином питної соди
Пріонові інфекції
Стимулюють утворення антитіл
Порушують функції клітинних мембран
Індукують продукцію інтерферону
Пріонові інфекції НЕ передаються
Повітряно-крапельним шляхом
Аліментарним шляхом
Трансфузіонним шляхом
Комахами
Виникнення губчастих енцефалопатій НЕ пов’язано з
Мутацією гену PrP
Поглинанням пріонових білків з їжею
Трансдукцією за допомогою вірусів
Віруси НЕ мають власних
Систем біосинтезу білків та виробництва енергії
Ферментів
Двонитчастих РНК
Метілювання вірусних геномів забезпечує
Початок транскрипції
Захист від клітинних рестриктаз
Позначає кінцеву ділянку транскріпції
Віроїди розмножуються
Автокатилізом
Поділом навпіл
Шляхом спороутворення
Пріони являють собою
Нуклеопротеїд
Глікопротеїд
Ланцюг з амінокислотних залишків
Пріони є збудниками
гострих респіраторних захворювань корів
губчастих енцефалопатій людини та тварин
хвороб кишково-щлункового тракту людини
Пріони інактивуються
Ультрафіолетовим опроміненням на протязі 15 хвилин
хлораміном на протязв 5 хвилин
Автоклавуванням при 136 градусах на протязі 18 хвилин /
2% розчином питної соди
Пріонові інфекції
Стимулюють утворення антитіл
Порушують функції клітинних мембран
Індукують продукцію інтерферону
Пріонові інфекції НЕ передаються
Повітряно-крапельним шляхом
Аліментарним шляхом
Трансфузіонним шляхом
Комахами
Виникнення губчастих енцефалопатій НЕ пов’язано з
Мутацією гену PrP
Поглинанням пріонових білків з їжею
Трансдукцією за допомогою вірусів
Віроїди розмножуються
Автокатилізом
Поділом навпіл
Шляхом спороутворення
Одноланцюгові, некодуючі послідовності РНК, що викликають захворювання рослин, мають назву
віроїди
віруси-сателіти
віруси
віріони
Віроїди у клітинах розмножуються
автокаталізом та з використанням ферментніи систем клітин
за допомогою ферментів клітин
абіогенним синтезом
поділом навпіл
брунькуванням
Віруси НЕ мають власних
Систем біосинтезу білків та виробництва енергії
Ферментів
Двонитчастих РНК
Метілювання вірусних геномів забезпечує
Початок транскрипції
Захист від клітинних рестриктаз
Позначає кінцеву ділянку транскріпції
Капсид з ув'язненою в ньому нуклеїновою кислотою віруса – геномом має назву
нуклеокапсид
суперкапсид
капсид
нуклеоїд
Пріонові інфекції НЕ передаються
повітряно-крапельним шляхом
половим шляхом
аліментарним шляхом
ентеральним шляхом
вертикальним шляхом
Пріони інактивуються
2 % розчином питної соди
автоклавуванням при 120 градусах протягом 1,5 годин
96 % етанолом
ультрафіолетовим опроміненням протягом 15 хвилин
розчином хлораміну
Виникнення губчастих енцефалопатій НЕ пов’язано з
трансдукцією пріонів за допомогою вірусів
хворобами вірусного походження
вакуолізацією нейронів
поглинанням пріонових білків з їжею
N- мутацією гена Pr
Пріонові білки відрізняються від нормальних клітинних білків
амінокислотною послідовністю
зменшеним відсотком α-спиральних структур
збільшеним відсотком β-складчастих структур
нечутливістю до дії протеаз
здатністю вікликати імунну відповідю в ураженому організмі
Віроїди розмножуються
Автокатилізом
Поділом навпіл
Шляхом спороутворення
Пріони являють собою
Нуклеопротеїд
Глікопротеїд
Ланцюг з амінокислотних залишків
Пріони є збудниками
гострих респіраторних захворювань корів
губчастих енцефалопатій людини та тварин
хвороб кишково-щлункового тракту людини
Збудники губчастих енцефалопатій являють собою
білок
глікопротеїд
нуклеопротеїд
ланцюг із ліпопротеінів
Терміном “віріон” позначається
N- заключна форма розвитку вірусу, яку він набуває виходячи із клітини
зовнішня оболонка вірусу з антигенними комплексами
нуклеїнова кислота вірусу в стані, інтегрованому з геномом клітини
цитоплазматична нуклеїнова кислота
Лабораторна прижиттєва діагностика губчастих енцефалопатій проводиться за допомогою
тесту на новонароджених мишах
імуно-ферментного аналізу
мікроскопії зрізів головного мозку
полімеразної ланцюгової реакцією
Д.І. Івановський експериментально встановив основні властивості вірусів, такі як
ультрамікроскопічні розміри
здатність до розмноження
репродукцію виключно у живиж організмах
здатність до росту на синтетичних середовищах
мінливість вірусів у процесі культивування
Едвардом Дженером була запропонована і впроваджена дієва вакцина проти
натуральної віспи
сказу
грипу
герпесу
Вакцина та вакцинація проти сказу була запропонована
Луі Пастером
Едвардом Дженером
Ілією Мечниковим
Джоном Ридом
У вірусів відсутні власні
системи біосинтезу білків
системи виробництва енергії
ферменти
дволанцюгові РНК.
сполуки, відмінні від білків та нуклеїнових кислот
До простих, мінімальних вірусів відносяться
вірус гепатиту А
вірус поліоміеліту
вірус мозаїчної хвороби тютюну
вірус грипу птахів
вірус грипу людини
вірус простого герпесу
Віруси, що міcтять дволанцюгову ДНК, за класифікацією Балтімора відносяться до типу
N- I
II
III
IV
Віруси, що містять одноланцюгову ДНК, за класифікацією Балтімора відносяться до типу
ІІ
І
III
IV
Віруси, що містять двохланцюгову РНК, за класифікацією Балтімора відносяться до типу
ІІІ
І
V
IV
Віруси, що містять одноланцюгову (+) РНК, за класифікацією Балтімора відносяться до типу
IV
І
V
ІІІ
Віруси, що містять одноланцюгову (-) РНК, за класифікацією Балтімора відносяться до типу
V
І
ІІ
VІ
Віруси, що містять двохланцюгову ДНК та реплікуються через стадію одноланцюгової РНК, за класифікацією Балтімора відносяться до типу
VІІ
І
ІІ
VІ
Subviral Agents (підвірусні агенти) – це
- віруси - сателіти
віроїди
пріони
фрагменти вірусних геномів
Віруси що містять одноланцюгову молекулу РНК та мають у своєму життєвому циклі стадію синтезу ДНК на матриці РНК – стадію зворотної транскрипції, за класифікацією Балтімора відносяться до типу
VІ
ІІ
ІІІ
VІ
До типу І за класифікацією Балтімора відносяться слідуючі віруси родин
Herpesviridae
Poxviridae
Adenoviridae
Papovaviridae
Papillomaviridae
Coronaviridae
До типу V за класифікацією Балтімора відносяться слідуючі віруси родин
Paramyxoviridae
Rhabdoviridae
Orthomyxoviridae
Papillomaviridae
Coronaviridae
До типу VII за класифікацією Балтімора відносяться слідуючі віруси родин
Hepadnaviridae
Rhabdoviridae
Orthomyxoviridae
Papillomaviridae
Зовнішня (суперкапсидна) оболонка вірусів складається з
білків клітинного походження
білків вірусного походження
глікопротеїдів
ліпопротеїдів
нуклеопротеїдів
Можливість абіогенного синтезу полімерів
була практично доказана
є гіпотезою
теоретично не можлива
практично не встановлена
Молекули РНК, здатні до каталітичної активності, отримали назву
рибозими
РНК-ази
риболази
рибози
Недоліками гіпотези абіогенного синтезу РНК є необхідність у продуктах, утворення яких у безклітинному середовищі не доказано, а саме
залишків фосфорної кислоти
рибози
вільних нуклеотидів
тіміну
РНК-полімерази
Утворення вірусів, згідно сучасних поглядів, можливе
як біогенним, так і абіогенним шляхом
за рахунок гранічного пристосування клітин до паразитизму
з клітинних органел, які мають власні геноми
за рахунок занесення з інших планет
В.М. Жданов вважав віруси «переносниками передового досвіду в біосфері» на підставі
можливості індукувати перенесення і рекомбінацію клітинного генетичного матеріалу
високої мутаційної мінливості вірусів
здатності вірусів до розмноження в організмах різних видів
наявності у вірусів фрагментованих геномів
До біологічних властивостей пріонових білків відносяться
відсутність екліпс-фази
тривалий інкубаційний період (від місяців до десятків років)
хронічна прогресуюча патологія (повільна інфекція)
відсутність запальних реакцій
відсутність гену, якій кодує синтез пріонового білку
До біологічних властивостей пріонових білків відносяться
нечутливість до дії інтерферону
не викликають імунної відповіді
відсутність патогенної дії на клітини in vitro
хронічна прогресуюча патологія (повільна інфекція);
зміна кола хазяїв при пасивуванні
наявність вірусів – переносників пріонових білків
До біологічних властивостей пріонових білків відносяться
відсутність антигенності
розмноження автокаталітичним шляхом
відтворення як в клітині, так і в штучній, безклітинній системі автокаталітичним шляхом
реплікація до титрів 100000 -100000000000 в мозку
зміна кола хазяїв при пасивуванні
стійкість до високих доз ультрафіолетового опромінення
Віруси відмінні від клітинніх форм за слідуючими ознаками
індукують у уражених клітинах синтез інтерферону
ростуть і розмножуються на мінімальному середовищі
здатні до бінарного поділу
чутливі до дії антибактеріальних препаратів
Геном вірусів ізольований від зовнішнього середовища
білковою оболонкою
ліпідною мембраною
гліколіпідами
нуклеопротеїдами
До вірусів з кубічною (ікосаедричною) симетрією відносяться
Iridovirus
Herpesvirus
Adenovirus
Picornaviridae
Papallomavirus
Rhabdovirus
До вірусів з спіральним (паличковидним) типом симетрії відноситься
вірус мозаїчної хвороби тютюну
вірус генітального герпесу
N-вірус імунодефіциту людини
вірус Т2
Мутаційна мінливість вірусів може бути обумовлена
змінами нуклеотидних послідовностей у геномі під час транскрипції
помилками під час трансляції.
системами репарації клітини
рекомбінацією між вірусними геномами поза клітиною
Внутрішніми джерелами формування генофондів вірусних популяцій НЕ є
включення у геном вірусу генетичного матеріалу клітини
рекомбінації між вірусними геномами, що знаходяться у одній клітині.
мутації
робота систем репарації ДНК
У вірусів відсутні власні
системи біосинтезу білків та виробництва енергії
ферменти
дволанцюгові РНК.
сполуки, відмінних від білків та нуклеїнових кислот
Мінливість вірусів грипу людини забезпечується, головним чином
рекомбінаціями між вірусами грипу різних типів
здобуттям вірусами ознак клітини-господаря
помилками під час трансляції вірусних білків
транслокацією генів у геномі вірусу
За допомогою бактеріофагів відбувається явище
трансдукції
трансформації
кон’югації
рестрикції
Метилювання вірусних геномів забезпечує
захист від клітинних рестриктаз
визначає для рибосоми початок трансляції
позначає кінцеву ділянку транскрипції
позначає початок транскрипції
Запрограмований зсув рамки зчитування під час транскрипції іРНК з вірусних ДНК геномів забезпечує
збільшення кількості трансльованих білків вірусного походження
мутації вірусів
послідовний синтез вірусних складових
інтеграцію вірусних та клітинних геномів
Зміна властивостей клітини (організму) під впливом явища трансдукції має назву
конверсія
трансфекція
розбудова
перевтілення
Явище трансдукції забезпечується участю
помірних бактеріофагів
ферментів цитолізу
віроїдів
вірулентних бактеріофагів
Явище трансдукції, під час якого переносятся завжди одні і тиж самі ділянки клітинного геному, має назву
специфічна трансдукція
послідовна трансдукція
неспецифічна трансдукція
банальна трансдукція
Мутаційна мінливість у представників родини ретровірусів (зокрема, ВІЛ) зумовлена головним чином
помилками зворотної транскриптази під час синтезу ДНК
включенням у геном вірусів фрагментів клітинного геному
системами репарації клітини-господаря
генетичною рекомбінацією між представниками різних родин вірусів
В ході специфічної трансдукції бактеріофаг лямбда забезпечує перенос слідуючих генів кишкової палички
gal, bio
pro, tir
lac, trp
exe, doc
РНК вірусів, яка є інформаційною, позначається як
N- «+»
«-»
«:»
«х»
Транскрипція геномів РНК вірусів відбувається за допомогою
РНК - залежних РНК полімераз
РНК - залежних ДНК полімераз
ДНК - залежних РНК полімераз
ДНК - залежних ДНК полімераз
Геном ДНК-вмісних вірусів може бути
N- дволанцюговим
N- одноланцюговим
лінійним
кільцевим
фрагментованим
Перевагою кільцевої структури ДНК вірусів перед лінійною є
стійкість молекули до дії клітинних нуклеаз
компактність упаковки
легкість реплікації
стійкість до дії противірусних препаратів
Присутність інвертованих нуклеотидних послідовностей на кінцях молекул ДНК вірусів забезпечує
здатність молекули замикатися у кільце
стійкість до нуклеаз
чутливість до інтерферону
відмінність вірусів один від одного
Геном РНК вірусів може бути
одноланцюговим
дволанцюговим
лінійним
кільцевим
фрагментованим
цілком складатися з нетранслюючих нуклеотидних послідовностей
Назву «неструктурні білки вірусів» мають білки, що
виявляються в зараженій клітині під час вірусної інфекції
не входять до складу віріону
забезпечують внутрішньоклітинну репродукцію вірусів на різних етапах
забезпечують реплікацію вірусних геномів
забезпечують захіст вірусного геному від зовнішнього середовища
Білки суперкапсидів вірусів представлені
глікопротеідами
нуклеопротеідами
ліпопротеідами
гліколіпідами
фосфопротеїнами
Глікопротеіди на поверхні суперкапсиду складних вірусів беруть участь у
адсорбції вірусів на поверхні клітини
транспорті віруса у клітину
реплікації віруса
виході вірусу з ураженої клітини
Глікопротеіди вірусів використовують у практиці для отримання
противірусних вакцин
сироваток
противірусних лікарських препаратів
протипаразитарних препаратів
Ліпіди у складних вірусів виявляються лише у складі
ліпопротеідної оболонки
запасних поживних речовин вірусів
вірусних ферментів
вірусних ліпаз
Популяційну структуру вірусів і характер процесів визначають слідуючі фактори
велика чисельність популяції
швидка зміна поколінь (в порівнянні з життєвим циклом організмів-господарів)
гаплоїдність та безстатевий спосіб розмноження
мала місткість геномів та відсутність генів, що повторюються
безперервність в динаміці епідемічногопроцесу
Фактори, що спричиняють зміни вірусних геномів
мутації
рекомбінації
включення в геном генетичного матеріалу клітини-хозяїна
потік генів
репарація геномів
Мутації забезпечують у вірусів
появу генів з новими функціями
перерозподіл генетичного матеріалу
утворення популяцій, поєднуючих властивості вихідних (батьківських) форм
утворення в популяціях однакових за генотипом форм
Антигенний дрейф у вірусів грипу (зміни структури поверхневих антигенів) обумовлюють
точкові мутації генів гемаглютиніну і нейрамінідази
точкові мутації гену гемаглютиніну
точкові мутації гену нейрамінідази
рекомбінації між фрагментами геному
Фермент, який утворює ланцюг ДНК на матриці РНК має назву
РНК-залежна ДНК-полімераза
зворотна транскриптаза
ревертаза
ДНК-полімераза
РНК- транскриптаза
Явище, при якому інфікування одним вірусом клітини повністю блокує можливість реплікації вірусу іншого виду в межах цієї клітини, має назву
гетерологічна інтерференція
гомологічна інтерференція
сімбіотична інтерференція
конверсія
При гомологічній інтерференції реплікація віруса з дефектним геномом можлива при
сумісному зараженні з нормальним вірусом
роботі системи репарації клітини
за допомогою клітинних полімераз
за допомогою зворотної транскрипції
Геном вірусів включає:
структурні гени
регуляторні гени
некодуючі ділянки (сайти)
ділянка attP, що забезпечує інтеграцію вірусного геному в хромосому клітини-хазяїна
ділянки cos - липкі кінцеві ділянки лінійних вірусних хромосом, що забезпечують замикання лінійної хромосоми в кільцеву форму
гени, що кодують рРНК і тРНК
РНК-геномні віруси, окрім ……….розмножуються в цитоплазмі
вірусу грипу і ретровірусів
параміксовірусів
вірусів сказу
вірусів поліоміеліту
Усі ДНК-вмісні віруси, окрім ………., розмножуються в ядрі
вірусу віспи
аденовірусів
вірусів герпесу
вірусів гепатиту В
Віруси, здатні викликати вірогенію (лізогенію), відносяться до групи
помірних
вірулентних
дефектних
незалежних
В основі неспецифічної адсорбції лежить механізм
Ван-дер-Ваальсових сил
закону Авогадро
закону Ньютона
теорії Ломоносова – Лавуазьє
В основі механізму специфічної адсорбції лежить
взаємодія між рецепторами вірусу та клітини
взаємодія між протилежними електричними зарядами
взаємодія між іРНК та рібосомою
взаємодія білків з нуклеїновими кислотами
На ефективність адсорбції вірусу на клітинній мембрані впливає
N- концентрація вірусів
температура
стан клітини
наявність в середовищі електролітів (катіонів) і вільних амінокислот-кофакторів
тканинна специфічність
У рецептосомі (при рецепторному піноцетозі) відбувається взаємодія
поверхневих білків віріонів з ліпідами стінки вакуолі
поверхневих ліпідів віріонів з ліпідами стінки
поверхневих білків віріонів з білками стінки
нуклеопротеідів віріонів з ліпідами стінки
Злиття поверхневих білків віріонів з ліпідами стінки вакуолі призводить до
виходу внутрішнього компонента вірусів в цитоплазму
інактивації віруса
реплікації віруса
депротеінізації віріону
Взаємодія вірусної і клітинної мембран відбувається в більшості випадків при наступних значеннях рН
при низьких (5,0 - 5,5)
при низьких (2,5 – 3,5)
при середніх (6,0 – 6,5)
при високих (8,0 – 9,0)
Інфікування вірусами бактерій реалізується. . . . . . . . . . шляхом
трансфузійним
аліментарним
повітряно-краплинним
статевим
Інформація, записана у вірусному геномі, у різних вірусів може реалізуватися шляхом
перемикання клітини на синтез вірусспецифічних молекул
утворенням коротких інформаційних РНК для індивідуальних білків
використанням вірусом власних геномів як інформаційних РНК
посттрансляційного нарізання білків
трансляції амінокислотної послідовності в нуклеотидну послідовність
Компоненти вірусу синтезуються в клітині
окремо
в одному місті
одночасно
залежно один від одного
Експресія вірусних геномів регулюється на рівні
транскрипції
трансляції
процесінгу
посттрансляційної репарації
Реплікація вірусів з двох ланцюговим (дл) ДНК геномом іде шляхом
(дл) ДНК – іРНК – Білок
(дл) ДНК –(ол) ДНК – іРНК – Білок
(дл) ДНК – рРНК - Білок
(дл) ДНК – тРНК – Білок
Реплікація вірусів з одноланцюговим (ол) ДНК геномом іде шляхом
(ол) ДНК – (дл) ДНК – іРНК – Білок
(ол) ДНК – іРНК – Білок
(ол) ДНК – рРНК - Білок
(ол) ДНК – тРНК – Білок
Реплікація вірусів з дволанцюговими (дл) РНК геномом іде шляхом
(дл) РНК – РНК/ДНК – ДНК/ДНК - іРНК – Білок
(дл) РНК - іРНК – Білок
(дл) РНК–(ол) ДНК - іРНК – Білок
(дл) РНК - тРНК – Білок
Реплікація вірусів з (+)РНК геномом іде шляхом
(+)РНК - Білок
(+)РНК – (-)РНК - Білок
(+)РНК – ДНК - Білок
(+)РНК – (-)РНК – Білок
Реплікація вірусів з (-)РНК геномом іде шляхом
(-)РНК – (+РНК) - Білок
(-)РНК – (+)РНК – ДНК - Білок
(-)РНК – ДНК - Білок
(-)РНК – ДНК – іРНК – Білок
Лімітуючим фактором у формуванні зрілих вірусних часток є
кількість та функціональна активність матриксного білку
ферментативна активність вірусних білків
активність вірусних лігаз
активність клітинних лігаз
Віруси рослин можуть переходити з інфікованої клітини в незаражену
через плазмодесми
шляхом «брунькування»
шляхом «вибуху»
за допомогою формування вакуолі у клітині
Віруси герпесу та респіраторно-синцитіальні віруси можуть проникати в незаражену клітину з інфікованої
через плазмодесми
шляхом «брунькування»
шляхом «вибуху»
за допомогою формування вакуолі у клітині
Вірус, що передається від одного покоління клітин до іншого в ході клітинного поділу в формі латентного вірусного геному, називається
провірусом або, у разі вірусів бактерій, профагом
віроідом
віріоном
вірогеном
Короткий (1 - 3 дні) інкубаційний період буває звичайно в тих випадках, коли вірус розмножується в клітинах
близьких до вхідних воріт інфекції
нервової системи
епітелію
лімфатичної системи
Повітряно-крапельним шляхом можуть передаватися віруси родин
Orthomyxoviridae
Hepadnaviridae
Retroviridae
Poxviridae
Аліментарним шляхом можуть передаватися віруси родин
Picornaviridae
Paramyxoviridae
Retroviridae
Hepadnavirida
Сатевим шляхом можуть передаватися віруси родин
Hepadnaviridae
Paramyxoviridae
Picornaviridae
Orthomyxoviridae
Ентеральним (трансфузійним) шляхом можуть передаватися віруси родин
Retroviridae
Orthomyxoviridae
Picornaviridae
Paramyxoviridae
Вертикальним шляхом можуть передаватися віруси родин
Retroviridae
Picornaviridae
Paramyxoviridae
Orthomyxoviridae
Для доказу того, що захворювання викликається певним вірусом, необхідно виконати так звані постулати
Ріверса
Коха
Пастера
Мечникова
Для доказу того, що захворювання викликається певним вірусом, необхідно виконати так звані постулати Ріверса
Виділити вірус з організму хворого
Культивувати вірус в організмі або в клітинах експеріментальної тварини
Довести фільтрованість інфекційного агента (щоб виключити бактерії)
Відтворити подібне захворювання у представника даного або близького виду
Повторно виділити той самий вірус
Синтез інтерферонів може бути реакцією клітини на
інфікування вірусом
етапом клітинної трансформації
проміжною данкою в процесах дихання
етапом біосинтезу ферментів
Індукторами інтерферону можуть бути
N- РНК віруси
ДНК віруси
бактеріальні ендотоксини
синтетичні полімери
лектини рослин
Метод виявлення специфічних включень в ураженій клітині має назву
вірусоскопічний
серологічний
вірусологічний
молекулярно-гібрідологічний
Найголовнішою перевагою перещеплюваних культур перед первинними є
потенційне безсмерття
гетероплоїдний каріотип
більш висока чутливість до одних і тих самих вірусів
чутливість до більш широкого кола вірусів
Реакція гемаглютинації
проводиться у полістиролових планшетах при безпосередньому контакті вірусних антигенів з еритроцитами
проводиться у дослідах на лабораторних тваринах
проводиться на культурах клітин, заражених гемаглютинуючими вірусами
проводиться на курячих ембріонах
Реакція нейтралізації і гальмування гемадсорбції застосовується для
виявлення вірусних часток безпосередньо у досліджуваному матеріалі за допомогою специфічної сироватки
визначення виду вірусу та його титру за допомогою відомої специфічної сироватки у культурі клітин
визначення вмісту противірусних антитіл у досліджуваній сироватці за відомою титру вірусу у культурі клітин
визначення титру антитіл безпосередньо у досліджуваній сироватці за допомогою стандартного діагностикуму з відомим титром вірусу
Патологічні мітози класифікують за
морфологічними ознаками дегенеративних змін у моношарі клітин
характером пошкоджень клітинних структур
механізмом виникнення патології мітозу
морфологічними ознаками та цитофізіологічними механізмами їх виникнення
Використання курячих ембріонів для культивування вірусів забезпечує
cтерильність середовища для культивування, наочність змін, що виникають, під дією вірусів
можливість диференціювати віруси птахів від вірусів тварин
можливість культивувати виключно віруси курей
можливість диференціювати віруси пташиного грипу від вірусів грипу людини
Реакція гемадсорбції – це
здатність клітин, інфікованих вірусом, адсорбувати на своїй поверхні еритроцити
здатність клітин, інфікованих вірусом, адсорбувати лейкоцити
здатність клітин, інфікованих вірусом, адсорбувати тромбоцити
здатність клітин, інфікованих вірусом, адсорбувати білок-гемоглобін
Титрування виділених вірусів проводять з метою
кількісного визначення вмісту вірусних часток в одиниці об’єму досліджуваного матеріалу
визначення видової приналежності вірусу
виділення вірусів
дослідження морфології вірусів
Імуно-ферментний аналіз (ІФА) – імунологічна реакції між
антигенами вірусу і гомологічними антитілами
антигенами різних вірусів
антитілами різних класів
білками вірусів
Реакція гемаглютинації – феномен зклеювання
еритроцитів
лейкоцитів
лімфоцитів
тромбоцитів
Реакція гальмування гемаглютинації (РГГ) –серологічна реакція, що застосовується для
ідентифікації вірусів
визначення рівня патогенності вірусів
титрування вірусів
визначення хімічного складу вірусів
До серологічного методу діагностики вірусних захворювань, зокрема, відноситься метод
імуно-ферментного аналізу
реакції гемаглютинації
полімеразної ланцюгової реакції
вірусоскопії
Метод виділення вірусів на іонних сорбентах грунтується на
наявності електричного заряду у вірусних часток
виявленні окремих амінокислот, білків вірусів
нуклеотидній послідовності вірусних геномів
ферментативних властивостях вірусних білків
Обробка вірусів із суперкапсидними оболонками (складних вірусів) хлороформом або ефіром призводить до
руйнування вірусів
концентрації вірусів
підвищенню рівня патогенності вірусів
фрагментації вірусних геномів
Вірусоскопічний метод полягае у
виявленні специфічних включень у ураженій клітині
виявленні специфічних клінічних ознак захворювання
виявленні специфічних для данного віруса нуклеотидних послідовностей
виявленні противірусних антитіл
Використання курячих ембріонів для культивування вірусів забезпечує
cтерильність середовища для культивування, наочність змін, що виникають під дією вірусів
можливість диференціювати віруси птахів від вірусів тварин
можливість культивувати виключно віруси курей
можливість диференціювати віруси пташиного грипу від вірусів грипу людини
Реакція гемадсорбції – це
здатність клітин, інфікованих вірусом, адсорбувати на своїй поверхні еритроцити
здатність клітин, інфікованих вірусом, адсорбувати лейкоцити
здатність клітин, інфікованих вірусом, адсорбувати білок-гемоглобін
Титрування виділених вірусів проводять з метою
кількісного визначення вмісту вірусних часток в одиниці об’єму досліджуваного матеріалу
визначення видової приналежності вірусу
виділення вірусів
дослідження морфології вірусів
Ростові середовища для культивування клітинних культур містять
амінокислоти, вітаміни, мінеральні солі, вуглеводи
амінокислоти, вітаміни, сироватку крові ссавців
амінокислоти, вітаміни, вуглеводи, сироватку крові ссавців
вітаміни, мінеральні солі, вуглеводи, сироватку крові ссавців.
Назвіть основні ознаки, які є характерними для патогенезу грипу
повітряно-краплинний шлях передачі
висока контагіозність
вірусемія
розвиток інтоксикацій
наявність вторинної бактеріальної інфекції
аліментарний шлях передачі
Основним неспецифічним фактором захисту організму від вірусів грипу є
інтерферон
вакцинація
антибіотики
активність Т-лімфоцитів
До обов’язкових хімічних компонентів вірусів грипу належать
білки
вуглеводи
нуклеїнова кислота
ліпіди
вітаміни
Найбільш енергетично вигідною структурою при кубічній симетрії вірусної частки є
ікосаедр
октаедр
тетраедр
додекаедр
Вірус грипу належить до родини
Orthomyxoviridae
Picornaviridae
Togaviridae
Retroviridae
Вірус імунодефіциту людини належить до родини
Retroviridae
Rhabdoviridae
Paramyxoviridae
Filoviridae
Вірус Ебола належить до родини
Filoviridae
Picornaviridae
Togaviridae
Retroviridae
Вірус простого герпесу належить до родини
Herpesviridae
Filoviridae
Poxviridae
Herpesviridae
Вірус гепатиту В належить до родини:
Hepadnaviridae
Filoviridae
Poxviridae
Herpesviridae
Які з поданих хвороб є пріонової етіології?
хвороба Крейтцфельда-Якоба
синдром Тернера-Шерешевського
скрепі
куру
коклюш
Основним лабораторним методом виділення і культивування вірусів простого герпесу є
зараженням хоріон-алантоїсної оболонки курячого ембріону
зараженням первинних клітинних культур
зараженням перещеплюваних клітинних культур
зараженням лвбораторних тварин
Вірус гепатиту С відноситься до родини
Flaviviridae
Picornaviridae
Reoviridae
Retroviridae
Заміна типів гемаглютиніну і нейрамінідази (шифт) у вірусів грипу відбувається внаслідок
рекомбінації між фрагментами вірусних геномів
інверсії
точкових мутацій генів гемаглютиніну і нейрамінідази
делецій великих ділянок геному
Рефлекторная дуга вегетативной
нервной системы
В простой трехнейронной вегетативной рефлекторной дуге (рис.2), как и в соматической, выделяют те же три звена, а именно: рецепторное, образованное чувствительным (афферентным) нейроном, ассоциативное, представленное вставочным (ассоциативным) нейроном и эффекторное звено, образованное двигательным (эффекторным) нейроном, передающим возбуждение на тот или иной рабочий орган. Нейроны связаны между собой синапсами, в которых с помощью медиаторов происходит передача нервного импульса с одного нейрона на другой.
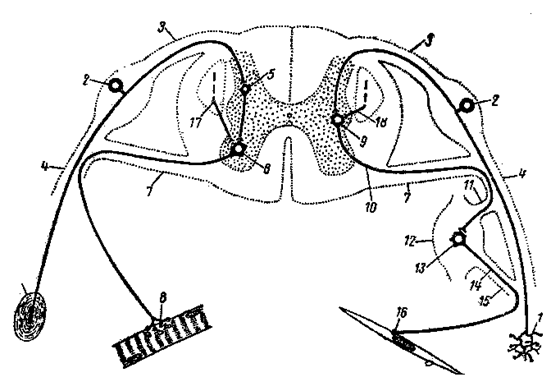
Рис. 2. Схема рефлекторных дуг соматического (слева) и вегетативного (справа) типов, замыкающихся в спинном мозге.
1- рецептор; 2- чувствительный нейрон спинномозгового ганглия; 3- дорсальный корешок; 4- спинномозговой нерв; 5- вставочный нейрон; 6- двигательный нейрон переднего рога; 7- вентральный корешок; 8- двигательное нервное окончание скелетной мышцы; 9- нейрон симпатического ядра бокового рога; 10- преганглионарное волокно; 11- белая соединительная ветвь; 12- периферический вегетативный ганглий; 13- эффекторный нейрон; 14- постганглионарное волокно; 15- серая соединительная ветвь; 16- двигательное нервное окончание на гладкой мышце; 17и 18- волокна пирамидного пути.
Чувствительные нейроны представлены псевдоуниполярными клетками спинномозгового узла, так же как и в соматической нервной системе. Их периферические отростки заканчиваются рецепторами в органах. Поэтому информация о состоянии органов растительной и животной жизни стекается в спинномозговые узлы, и в этом смысле они являются смешанными соматически-вегетативными узлами. Центральный отросток чувствительного нейрона в составе заднего корешка вступает в спинной мозг и нервный импульс переключается на вставочный нейрон, клеточное тело которого расположено в боковых рогах (латерально-промежуточное ядро тораколюмбального или сакрального отделов) серого вещества спинного мозга.
Аксон вставочного нейрона покидает спинной мозг в составе передних корешков и достигает одного из вегетативных узлов, где вступает в контакт с эффекторным (двигательным) нейроном.
Таким образом, второе звено вегетативной рефлекторной дуги отличается от соматической, во-первых, местом локализации тела вставочного нейрона, во-вторых, протяженностью и положением аксона, который в отличие от соматической нервной системы никогда не остается в пределах спинного мозга. Еще большие различия в строении третьего звена рефлекторной дуги. В отличие от соматической рефлекторной дуги, где двигательные нейроны расположены в передних рогах спинного мозга, для вегетативной рефлекторной дуги характерно расположение двигательного нейрона за пределами центральной нервной системы - в вегетативных узлах, аксоны которых направляются к рабочему органу, а это значит, что весь эфферентный путь подразделяется на два участка: предузловой (преганглионарный) - аксон вставочного нейрона и послеузловой (постганглионарный) - аксон двигательного нейрона вегетативного узла. Таким образом, в вегетативной рефлекторной дуге эфферентный периферический путь двухнейронный.
В простой трехнейронной вегетативной рефлекторной дуге, замыкающейся в пределах мозгового ствола, тело первого нейрона располагается в чувствительных узлах черепных нервов, второго - в вегетативных ядрах черепных нервов (мезенцефалический и бульбарный отделы) и третьего - в вегетативных узлах.
Достигая эффекторов (гладких мышц и желез), нервные импульсы вызывают сокращение мышцы или изменение секреторной деятельности железы, что в свою очередь вызывает раздражение рецепторов этих органов и отсюда поток импульсов по афферентным волокнам направляется обратно в ядра спинного или головного мозга, неся ежемоментную информацию о состоянии данного органа. Наличие обратной связи (обратной афферентации), с одной стороны позволяет осуществлять контроль за правильностью исполнения команд, с другой - вносить дополнительную своевременную коррекцию в выполнении ответной реакции организма.
Таким образом, в основе строения и функции вегетативной нервной системы, как и соматической, лежит замкнутая кольцевая цепь рефлексов, которая способствует наиболее полному приспособлению организма к окружающей среде.
ВЕГЕТАТИВНЫЕ УЗЛЫ
Узлы вегетативной нервной системы устроены однотипно, но отличаются друг от друга своей локализацией. По этому топографическому признаку их делят условно на три группы (порядка).Узлы I порядка, околопозвоночные (паравертебральные), ganglia trunci sympathici, образуют симпатический ствол, расположенный по сторонам позвоночного столба. Узлы II порядка, предпозвоночные (превертебральные) или промежуточные, ganglia intermedia, расположены впереди позвоночника, входят в состав вегетативных сплетений и также как и узлы I порядка относятся к симпатическому отделу вегетативной нервной системы. Узлы III порядка составляют конечные (терминальные) узлы, ganglia terminalia. Они в свою очередь разделяются на околоорганные и внутриорганные и относятся к парасимпатическим узлам.
Различная степень удаленности вегетативных узлов от спинного мозга или мозгового ствола, с одной стороны, и рабочего органа, с другой, сказывается на длине преганглионарных и постганглионарных волокон. Наибольшую длину имеют преганглионарные волокна, связанные с узлами III порядка. Постганглионарные волокна этих узлов, наоборот, очень короткие. Обратная картина наблюдается при рассмотрении волокон, связанных с узлами I порядка. Здесь преганглионарные волокна короткие, а постганглионарные - длинные. Преганглионарные волокна покрыты миелиновой оболочкой, благодаря чему имеют белый цвет. Их диаметр чаще составляет 2-3,5 мкм, а скорость распространения импульсов колеблется от 3 до 18 м/сек. Постганглионарные волокна лишены миелина, серого цвета, диаметр - до 2 мкм, скорость проведения импульса - 1-3 м/сек.
Снаружи вегетативный узел покрыт соединительнотканной капсулой, от которой внутрь отходят тяжи, между которыми располагаются группы клеток. Тело каждого нейрона имеет тонкую соединительнотканную капсулу. Между ней и телом нейрона - клетки сателлиты, которые выполняют опорную, защитную и трофическую функции.
В узлах выделяют три типа нейронов:
1. Клетки Догеля первого типа - мультиполярные с множеством коротких ветвящихся дендритов и одним длинным аксоном, направляющимся к рабочему органу (рис. 3). Это двигательные нейроны.
2. Клетки Догеля второго типа - чувствительные, мультиполярные с длинными дендритами, которые покидают узел и заканчиваются рецепторами на гладкомышечных или железистых клетках. Аксон может заканчиваться здесь же в ганглии, или направляться в соседние.
3. Клетки Догеля третьего типа представляют местные ассоциативные элементы, соединяющие своими отростками несколько клеток I и II типов как своего узла, так и соседних узлов.

| Рис.3. Различные типы нейронов соматической (а) и вегетативной (б) частей нервной системы (из А.Г.Кнорре и И.Д.Лев). 1- пирамидная клетка коры головного мозга; 2- клетка Пуркинье из коры мозжечка; 3- клетка-зерно из коры мозжечка; 4- двигательная клетка переднего рога спинного мозга; 5- биполярная клетка сетчатки; 6- униполярная клетка сетчатки; 7- клетка 1 типа Догеля; 8- клетка II типа Догеля; 9- клетка промежуточно-латерального ядра бокового рога спинного мозга. |
Вегетативные узлы выполняют следующие функции:
1. Передачу нервного импульса с преганглионарной на постганглионарную часть эфферентного пути. Эта функция обусловлена наличием в узле двигательных нейронов.
2. Рефлекторную или замыкательную. Благодаря наличию не только двигательных , но и чувствительных нейронов в вегетативных узлах могут замыкаться периферические рефлекторные дуги, что превращает их в периферические нервные центры, способные осуществлять местные рефлексы на основе принципа саморегуляции.
3. Рецепторную, так как в узлах имеются рецепторы, которыми заканчиваются дендриты чувствительных нейронов спинномозговых и черепных узлов. Благодаря центростремительным связям состояние и деятельность вегетативных узлов находятся под контролем центральной нервной системы.
4. Через вегетативный узел проходят транзитно афферентные и эфферентные волокна.
Кроме анатомически обособленных ганглиев, по ходу вегетативных ветвей периферических нервов встречается большое количество нервных клеток, мигрировавших сюда в ходе эмбрионального развития.
МНОГОЭТАЖНАЯ СТРУКТУРА МЕХАНИЗМА
РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Чтобы лучше понять взаимоотношение отделов нервной системы, регулирующих деятельность внутренних органов обратимся к схеме, на которой можно выделить 6 “этажей” (рис. 4).
Каждый отдел или “этаж” связан с внутренними органами эфферентными и афферентными путями и здесь происходит замыкание рефлекторных дуг.
Первый “этаж”, т.е. периферический вегетативный узел, является единственным отделом, который получает непосредственную афферентную связь с
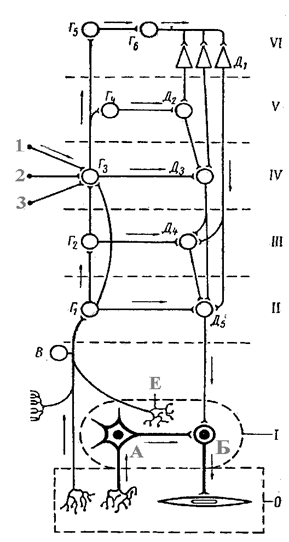
| Рис.4. Схема основных связей центральной и вегетативной нервной системы (по С.П.Семёнову). О- орган; I- периферический ганглий; II- сегмент спинного мозга или ствола; III- надсегментарные центры продолговатого мозга; IV- промежуточный мозг; V- ядра основания; VI- кора больших полушарий; А- чувствительный нейрон вегетативного ганглия; Б- эффекторный вегетативный нейрон; В- чувствительные нейроны спинномозговых и черепномозговых ганглиев; Г1 - Г4 - ассоциативные ядра разных отделов мозга; Г5 - клетки афферентных зон коры; Г6 - ассоциативные клетки; Д1 - пирамидные клетки коры; Д2 - Д5- вегетативные центры; Е- рецептор вегетативного ганглия;1-3 - проводящие пути из органов чувств. I - периферический ганглий; II - сегментарные центры спинного мозга и мозгового ствола; III - надсегментарные центры сетчатой формации; IV - промежуточный мозг; V - базальные ядра полушарий; VI - кора больших полушарий. |
внутренними органами, что предохраняет центральную нервную систему от потока избыточной информации, а также это единственный отдел, который не получает нервных импульсов непосредственно из коры больших полушарий.
Периферические рефлекторные дуги могут замыкаться в узлах симпатического ствола, узлах сплетений и интраорганных (интрамуральных) узлах.
Чем короче рефлекторная дуга, чем ниже её уровень, тем выше степень её функциональной автономии, в связи с чем интрамуральные ганглии обладают значительно большей степенью функциональной автономии и выполняют роль интегративной системы. Строение интрамуральных ганглиев больше напоминает строение центральной нервной системы, чем пре- и паравертебральные ганглии (отсутствие соединительной ткани, изоляция от кровеносных сосудов, незначительные внеклеточные пространства, многочисленные глиальные элементы, плотный синаптический нейропиль). В связи с вышеизложенным в настоящее время кроме симпатического и парасимпатического отделов вегетативной нервной системы выделяют и метасимпатический или энтеральный отдел, в состав которого входит комплекс микроганглионарных образований, расположенных в стенках внутренних органов (ауэрбахово, мейснерово сплетение, микроганглии в толще сердечной мышцы, мочевом пузыре, матки, стенках бронхов и т.д.).
Метасимпатическая система рассматривается как независимая система, которая программирует и координирует двигательную активность и другие функции висцеральных органов, такие как секреция и локальный кровоток и способна функционировать при полной децентрализации.
МОРФО-ФУНКЦИОНАЛЬНЫЕ РАЗЛИЧИЯ
ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Вегетативная нервная система отличается от соматической следующими признаками:
1. Вегетативный отдел нервной системы иннервирует гладкую мускулатуру, которая сокращается непроизвольно (без контроля сознания), медленно, а в отдельных органах находится в состоянии длительного сокращения (сфинктеры полых органов) или постоянного сокращения (гладкие мышцы стенок кровеносных сосудов). Соматический отдел иннервирует скелетную мускулатуру и побуждает её к выполнению быстрых и целенаправленных сокращений, осуществляющихся под контролем сознания.
2. Кроме того, что вегетативный отдел нервной системы иннервирует гладкую мускулатуру внутренних органов и сосудов, мышцу сердца и железы, она обеспечивает трофическую иннервацию всех тканей и органов, включая скелетную мускулатуру, т.е. отличается от соматического отдела повсеместностью распространения.
3. Важнейший отличительный признак вегетативного отдела - это очаговый характер расположения центров (ядер) в стволе головного мозга (мезенцефалический и бульбарный отделы) и спинном мозге (тораколюмбальный и сакральный отделы). Соматические же центры располагаются в пределах центральной нервной системы равномерно (сегментарно).
4. Очаговость расположения центров предопределяет и очаговый характер выхода корешков вегетативных волокон от мозгового ствола и спинного мозга. Они выходят в составе III, YII, IX, X пар черепных и C8 - L3, S2 - S4 спинномозговых нервов. При этом принцип строгой сегментарности отсутствует не только в месте отхождения, но и в распределении волокон на периферии.
5. Различия в строении рефлекторной дуги (см. выше).
6. Деятельность вегетативной нервной системы основана не только на центральных рефлекторных дугах, но и на периферических, двухнейронных, замыкающихся в вегетативных узлах.
7. Вегетативная нервная система сохранила ряд примитивных черт в своём строении:
а) диффузность локализации нейронов в виде ганглиев;
б) сравнительное однообразие форм и размеров нейронов, нервные клетки как правило мелкие, многоотростчатые (рис. 3);
в) меньший калибр нервных волокон из-за тонкой миелиновой оболочки (преганглионарные волокна) или ее отсутствия (постганглионарные волокна), что обусловливает низкую возбудимость и скорость проведения импульса. В соматических афферентных волокнах скорость составляет 70-120 м/сек, в вегетативных - 0,3-10 м/сек.
8. Вегетативная нервная система обладает избирательной чувствительностью к гормонам и различного рода метаболитам. Это обусловлено тем, что переключение импульса в синапсах осуществляется с помощью химического вещества - медиатора. Некоторые биологически активные вещества и гормоны являются стимуляторами в механизмах освобождения и выделения медиаторов и тем самым повышают тонус симпатического или парасимпатического отдела вегетативной нервной системы.
9. Для вегетативной нервной системы характерным является принцип многосегментарности в иннервации органов, что связано с процессами периферической и продольной миграции клеток в эмбриогенезе.
РАЗВИТИЕ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
ФИЛОГЕНЕЗ
Впервые намечается дифференцировка нервной системы на вегетативный и соматический отделы у ланцетника, у которого наблюдаются внутриорганные вегетативные сплетения. На этой стадии развития еще отсутствуют вегетативные узлы. У круглоротых (минога) и хрящевых рыб (акула, скат), помимо вегетативных внутриорганных сплетений, уже имеются блуждающие нервы, достигающие сердца и проксимального отдела пищеварительного тракта, появляются зачатки узлов будущих симпатических стволов в виде диффузных нервных сплетений по бокам аорты. Преганглионарные волокна выходят не только в составе вентральных корешков, но и дорсальных.
Симпатический ствол впервые появляется у костистых рыб. У амфибий происходит дальнейшая концентрация вегетативных нейронов, формирование экстраорганных сплетений и перераспределение вегетативных волокон в сторону их увеличения в составе вентрального корешка.
В ряду позвоночных расширяется территория распространения ветвей блуждающих нервов. У млекопитающих и человека происходит интенсивное развитие головных узлов вегетативной нервной системы, отмечается четкая дифференцировка клеток вегетативных узлов на 3 типа. С появлением головного мозга, а также коры головного мозга развиваются интегрирующие центры, берущие на себя роль высших (надсегментарных) центров вегетативной нервной системы.
Таким образом, в ряду позвоночных эволюция вегетативной нервной системы выражается в превращении диффузных сплетений в ганглии, а также переход преганглионарных волокон из дорсальных корешков в вентральные.
Итак, в процессе развития вегетативная нервная система специализируется в другом направлении, чем соматическая нервная система. Последняя эволюционирует быстрее, вегетативная же несколько отстает и сохраняет черты более примитивной нервной системы.
ЭМБРИОГЕНЕЗ
На 3-й неделе эмбрионального развития человека в головном и спинном мозге в результате размножения и дифференцировки нейробластов возникают вегетативные центры.
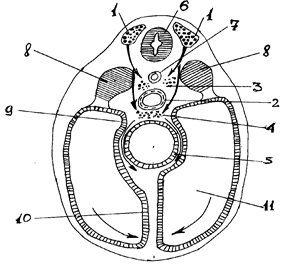
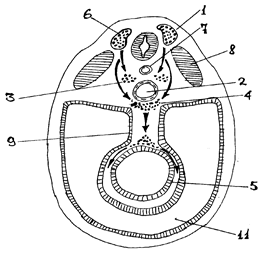
Источником нейронов периферического отдела вегетативной нервной системы является ганглиозная пластинка, которая возникает из нервных валиков (переходная область между нервной пластинкой и кожной эктодермой). Название “ганглиозной пластинки” обусловлено тем, что из нее образуются все нервные узлы. В основе образования вегетативных узлов лежит миграция клеток из гаглиозной пластинки на периферию (рис.5). Первыми закладываются узлы I порядка (околопозвоночные), несколько позже - узлы II (предпозвоночные) и III (органные) порядков. На первой стадии в результате периферической миграции клеток из ганглиозной пластинки образуются первичные сегментарные закладки вначале грудных и шейных, а затем, к концу I-го месяца, поясничных и крестцовых узлов симпатического ствола. На второй стадии происходит накопление клеток за счет прибывающих новых клеток, а также благодаря размножению уже прибывших. При этом происходит перемещение клеток вдоль симпатического ствола (продольная миграция) и первичные нервные узлы сливаются между собой, образуя сплошной несегментированный клеточный тяж (рис.6). На третьей стадии из клеточного тяжа формируются дефинитивные узлы симпатического ствола, которые, благодаря продольной миграции, содержат нейроны из многих сегментов ганглиозной пластинки.
| 
|
| Рис. 5. Схема нейрального пути миграции клеток ганглиозной пластинки (по П.И.Лобко). 1- ганглиозная пластинка; 2- аорта; 3- закладка симпатического ствола; 4- закладка предпозвоночных сплетений; 5- кишка; 6- нервная трубка; 7- спинная струна (хорда); 8- сомиты; 9- дорсальная брыжейка; 10- вентральная брыжейка; 11- вторичная полость тела (целом). |
Предпозвоночные и органные вегетативные узлы формируются за счет продолжающейся в вентральном направлении миграции клеток и их размножения (рис.6).
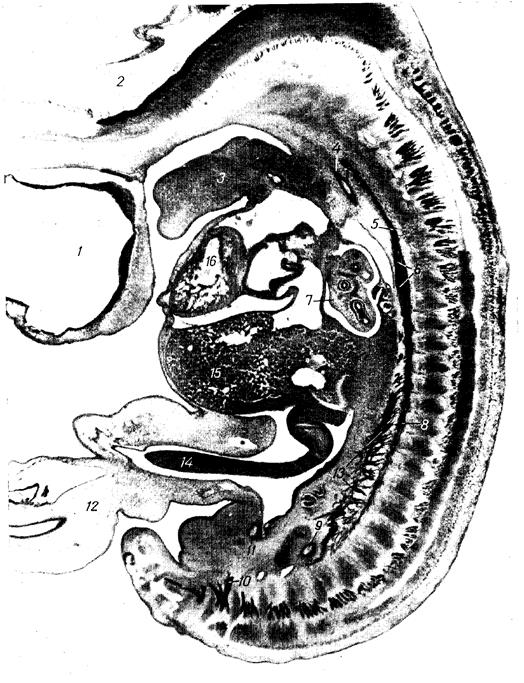
|
Рис. 6. Сагиттальный срез зародыша человека 12 мм длины (препарат Д.М.Голуба).
1- передний мозговой пузырь; 2- задний мозговой пузырь; 3- подъязычный нерв закладки языка; 4- правая дорсальная аорта; 5- шейный отдел симпатического ствола; 6- закладка звёздчатого узла; 7- лёгкое; 8- грудной отдел симпатического ствола; 9- каудальный отрезок нисходящей аоты; 10- третий крестцовый спинномозговой нерв; 11- пупочная артерия; 12- пупочный канатик; 13- висцеральные ветви симпатического ствола; 14- кишечная петля; 15- печень; 16- правый желудочек.
Нарушение миграционных процессов, лежащих в основе формирования вегетативных узлов, проявляется различными тяжелыми заболеваниями, например, врожденный аганглиоз толстой кишки (болезнь Гиршпрунга), мочевого пузыря и др. органов.
Нервные волокна различной функциональной принадлежности появляются неодновременно. Первыми обнаруживаются нервные волокна центрального происхождения - преганглионарные и афферентные, которые выходят из спинного и головного мозга в составе черепных и спинномозговых нервов. При этом преганглионарные волокна, вступающие в симпатический ствол, распространяются на значительные расстояния вверх и вниз и каждый узел получает их из многих сегментов. Такой характер распределения преганглионарных волокон, а также процесс продольной миграции клеток создают предпосылки к многосегментарной иннервации органов.
Исследования Д.М. Голуба и его сотрудников, выяснившие пути и последовательность врастания чувствительных нервных волокон в различные внутренние органы зародыша, позволили обосновать экспериментально на животных, а затем применить на больных людях в клинике ряд хирургических приемов восстановления чувствительной иннервации в тех органах, которые ее утратили в результате того или иного патологического процесса.
По мере дифференцировки нейронов вегетативных узлов появляются постганглионарные волокна, которые направляются к внутренностям, где образуют эфферентные окончания. Постганглионарные симпатические волокна, которые присоединяются к спинномозговым нервам (серые соединительные ветви) от одного узла симпатического ствола распределяются по многим спинномозговым нервам.
СИМПАТИЧЕСКИЙ И ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛЫ И ИХ РАЗЛИЧИЯ
На основе анатомических и функциональных различий в вегетативной нервной системе выделили два отдела - симпатический и парасимпатический.
Симпатический отдел по своим основным функциям является трофическим. Он обеспечивает усиление окислительных процессов, усиление дыхания, учащение деятельности сердца, т.е. приспосабливает организм к условиям интенсивной деятельности. В связи с этим тонус симпатической нервной системы преобладает днем, а ночью - парасимпатической (“царство вагуса”). Парасимпатический отдел выполняет охранительную роль (сужение зрачка, бронхов, снижение частоты сердечных сокращений, опорожнение полостных органов). Эффекты симпатического раздражения более диффузны и генерализованы, парасимпатического – более локальны.
Симпатический и парасимпатический отделы часто действуют как антагонисты. Однако антагонизм этот относительный. При резко измененном функциональном состоянии органа они могут действовать однонаправленно как синергисты. В ответ на усиленную деятельность организма, наступают и парасимпатические сдвиги, направленные на восстановление энергетического потенциала и гомеостаза. Благодаря активности и синергизму обоих отделов вегетативной нервной системы возможна длительная, адаптивная деятельность организма.
Таким образом, между ними существует не столько антагонизм, сколько взаимодействие, которое обеспечивает наиболее тонкое регулирование деятельности органов.
Следующее существенное отличие – различная медиация на окончаниях постганглионарных волокон (медиатором преганглионарных как симпатических так и парасимпатических волокон является ацетилхолин, действие которого потенцируется присуствием ионов калия). На окончаниях симпатических волокон выделяется симпатин (смесь адреналина и норадреналина), оказывающий местное влияние, а после всасывания в кровоток – общее. Медиатором парасимпатических постганглионарных волокон является ацетилхолин, который вызывает преимущественно местное воздействие и быстро разрушается холинэстеразой.
Наряду с функциональными имеются ряд морфологических различий симпатического и парасимпатического отделов вегетативной нервной системы, а именно:
1. Очаги выхода парасимпатических волокон из мозга разобщены друг от друга (мезенцефалический, бульбарный, крестцовый отделы), симпатические волокна выходят из одного, но более протяженного очага (тораколюмбальный отдел).
2. К симпатическим узлам относятся узлы I и II порядка, к парасимпатическим - III порядка (конечные). В связи с чем преганглионарные симпатические волокна более короткие, а постганглионарные более длинные, чем парасимпатические.
3. Сфера действия парасимпатической системы относительно ограничена и касается главным образом внутренних органов. Симпатический же отдел вегетативной нервной системы кроме внутренних органов иннервирует все сосуды, потовые, сальные железы и волосковые мышцы кожи, а также скелетную мускулатуру, обеспечивая ей трофическую иннервацию, то есть не существует каких-либо тканей, органов, систем куда бы не проникали симпатические волокна.
СИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ
Симпатическая нервная система состоит из центрального и периферического отделов.
Центральный отдел представлен ядрами боковых рогов серого вещества спинного мозга
Дата добавления: 2015-03-19; просмотров: 1493;