Виды эквивалентных электрических схем тканей организма
Электрические свойства биологических тканей моделируются сочетанием элементов, обладающих омическими и емкостными свойствами. Эти свойства можно моделировать с помощью эквивалентных электрических схем. Рассмотрим три из них, рис. 17.
Схема I не совсем удовлетворительна, т.к. содержит бесконечно большое сопротивление постоянному току (емкость не проводит постоянный ток), чего не наблюдается в тканях организма. Поэтому при низкой частоте схема I даст существенные отклонения расчетных значений импеданса (полного сопротивления) от реального импеданса биоткани. В схеме // при увеличении частоты тока ω емкостное сопротивление стремится к нулю 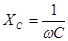 поэтому импеданс схемы также стремится к пулю. Это противоречит опыту.
поэтому импеданс схемы также стремится к пулю. Это противоречит опыту.
 У живых объектов импеданс уменьшается по мере увеличения частоты только до определенного значения. Для живых тканей характерно более сложное сочетание параллельного и последовательного соединений элементов, например, схема ///.
У живых объектов импеданс уменьшается по мере увеличения частоты только до определенного значения. Для живых тканей характерно более сложное сочетание параллельного и последовательного соединений элементов, например, схема ///.
В состоянии покоя реактивное сопротивление ткани носит только емкостный характер, при возбуждении ткань приобретает индуктивные свойства и обладает индуктивным сопротивлением
XС =ω L.
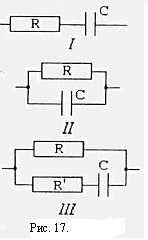
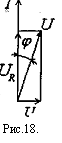 При анализе электрических свойств ткани рассматриваем только состояние покоя. Найдем импеданс Z простейшей эквивалентной электрической схемы /, рис. Векторная диаграмма имеет вид, показанный на рис.17. Общее напряжение равно
При анализе электрических свойств ткани рассматриваем только состояние покоя. Найдем импеданс Z простейшей эквивалентной электрической схемы /, рис. Векторная диаграмма имеет вид, показанный на рис.17. Общее напряжение равно
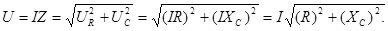
Следовательно, импеданс цепи равен
Угол сдвига фазы между током I и напряжением U найдем из условия.
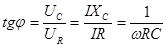
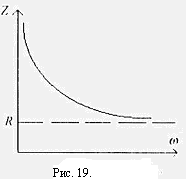 На рис.19. показана зависимость Z импеданса эквивалентной электрической схемы / (рис.17.) от частоты, построенная но формуле
На рис.19. показана зависимость Z импеданса эквивалентной электрической схемы / (рис.17.) от частоты, построенная но формуле 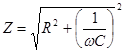
 Как видно из рис. 19., импеданс цепи монотонно падает с увеличением частоты электрического тока, постепенно приближаясь к активному сопротивлению R..
Как видно из рис. 19., импеданс цепи монотонно падает с увеличением частоты электрического тока, постепенно приближаясь к активному сопротивлению R..
Величины логарифмов импеданса и относительной диэлектрической проницаемости можно считать по модулю примерно пропорциональными друг другу.
Зависимость импеданса (или относительной диэлектрической проницаемости) биологических тканей от частоты электрического тока называется дисперсией импеданса (или относительной диэлектрической проницаемости), а график этой зависимости дисперсионной кривой.
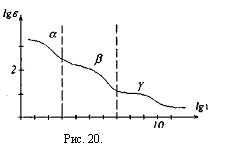
В отличие от зависимости импеданса от частоты электрического тока для эквивалентной электрической схемы биологической ткани, реальная дисперсионная кривая, например, скелетной мышцы, немонотонно падает с увеличением частоты, рис.20.
При действии на ткани организма переменного электрического тока или переменного электрического поля наблюдаются три области дисперсии. Область а - дисперсии.
Наблюдается до частот 1кГц (lgv≈3)
В низкочастотной части спектра области а- дисперсии электрический ток течет практически только по межклеточной жидкости, т.к. сопротивление мембран клеток очень велико. В высокочастотной части спектра импеданс падает за счет снижения емкостного сопротивления мембран клеток Хс = 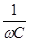 Возникает ток смещения, текущий через мембрану.
Возникает ток смещения, текущий через мембрану.
Область β- дисперсии.Наблюдается до частот I04 кГц (Igv≈3 – 7)
За счет увеличения частоты переполяризации мембран клеток снижается емкостное сопротивление ткани. Переполяризация рассматривается относительно пулевого уровня потенциала па мембране, за который принят потенциал покоя.
Затраты энергии на поляризацию и нереполяризацию клеточных структур (мембран, белковых молекул, молекул воды, атомов и т.д.) определяют импеданс ткани. Снижение этих затрат приводит к снижению импеданса биоткани.
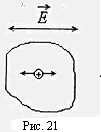 Многие ионы внутри клеток, рис.21, при такой частоте колебаний напряженности электрического поля
Многие ионы внутри клеток, рис.21, при такой частоте колебаний напряженности электрического поля  не успевают дойти до клеточных и внутриклеточных мембран. Для них сопротивление мембран роли не играет.
не успевают дойти до клеточных и внутриклеточных мембран. Для них сопротивление мембран роли не играет.
В высокочастотной области β- дисперсии существенное значение приобретает структурная поляризация и переполяризация крупных, в частности белковых молекул. Структурная поляризация заключается в том, что внутри молекулы, под действием электрического поля, смещаются ионы и ионные группы, входящие в состав молекул, что влияет на функциональные свойства белков.
Область γ дисперсии. Наблюдается до частот 107 кГц (Igv =• 7 - 10).
 За счет высокой частоты колебаний электрического тока, емкостное сопротивление мембран очень мало и они не представляют большого препятствия для переменного тока. Поэтому сопротивление биоткани в общем мало.
За счет высокой частоты колебаний электрического тока, емкостное сопротивление мембран очень мало и они не представляют большого препятствия для переменного тока. Поэтому сопротивление биоткани в общем мало.
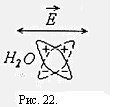
В низкочастотной части спектра области γ- дисперсии уменьшение диэлектрической проницаемости обусловлено постепенным ослаблением ориентационной переполяризации дипольных молекул воды, рис.22. В этой области важный вклад в импеданс ткани вносит структурная поляризация крупных молекул. Энергия электрического тока с увеличением частоты все меньше теряется на переориентацию молекул воды и структурную переполяризацию крупных молекул, поэтому уменьшается импеданс ткани.
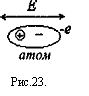
В высокочастотной части спектра γ - дисперсии молекулы воды под действием переменной напряженности Е почти не переполяризуются. Уровень электрического сопротивления определяется электронной переполяризацией атомов, т. с. возникновением переменных атомных диполей, рис.23.
Дата добавления: 2014-12-20; просмотров: 17281;